Рецептивное поле рефлекса это
В сетчатке человека имеются два типа ганглиозных клеток, отличающихся реакцией на точечные световые стимулы, воздействовавшие на центр или периферию их рецептивного поля (рис. 17.9). Примерно половина ганглиозных клеток возбуждается действием света на центр рецептивного поля и тормозится при действии светового стимула на периферию рецептивного поля. Такие клетки принято называть оn-нейронами. Другая половина ганглиозных клеток возбуждается действием светового раздражителя на периферию рецептивного поля и тормозится в ответ на световую стимуляцию центра рецептивного поля — они получили название off-нейронов.
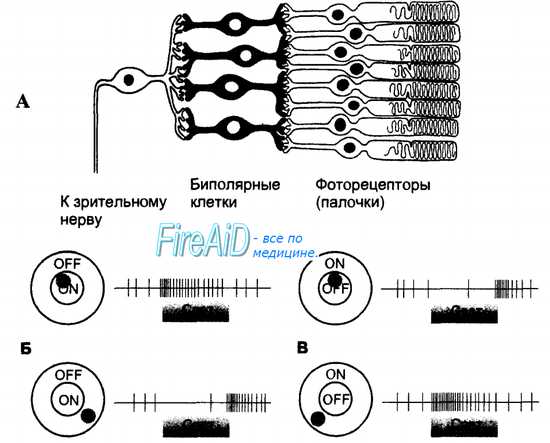 Рис. 17.9. Рецептивные поля ганглиозных клеток сетчатки с on- и off-центрами.
Рис. 17.9. Рецептивные поля ганглиозных клеток сетчатки с on- и off-центрами.
A. Рецептивное поле ганглиозной клетки образуют все фоторецепторы и биполярные клетки, имеющие с нею синаптические контакты. Ганглиозные клетки постоянно генерируют потенциалы действия, частота возникновения которых зависит от активности фоторецепторов и биполярных клеток, входящих в ее рецептивное поле.
Б. Ганглиозная клетка on-типа увеличивает частоту электрических разрядов в ответ на световое раздражение центра рецептивного поля и снижает свою электрическую активность при действии светового раздражителя на периферию рецептивного поля.
B. Ганглиозная клетка off-типа тормозится при действии света на центр ее рецептивного поля и увеличивает частоту нервных импульсов в ответ на раздражение периферии рецептивного поля.
Рецептивные поля ганглиозных клеток обоих типов в сетчатке представлены поровну, чередуясь друг с другом. Оба типа клеток очень слабо отвечают на равномерную диффузную засветку всего рецептивного поля, а наиболее сильным раздражителем для них является световой контраст, т. е. различная интенсивность засветки центра и периферии. Именно контрастирование деталей изображения дает необходимую информацию для зрительного восприятия в целом, тогда как абсолютная интенсивность отраженного от наблюдаемого объекта света не столь важна. Восприятие граней, т. е. восприятие контраста между соседними поверхностями с разной освещенностью, является наиболее информативным признаком изображения, определяющим протяженность и позиции разных объектов.
Рецептивное поле рефлекса это
Рецептивным полем называется область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 17.1). Максимальная величина рецептивного поля первичного сенсорного нейрона определяется пространством, которое занимают все ветви его периферического отростка, а число рецепторов, имеющихся в этом пространстве, указывает на плотность иннервации. Высокая плотность иннервации сочетается, как правило, с малыми размерами рецептивных полей и, соответственно, высоким пространственным разрешением, позволяющим различать стимулы, действующие на соседние рецептивные поля. Малые рецептивные поля типичны, например, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов значительно выше, чем на периферии сетчатки или в коже спины, для которых характерны большая величина рецептивных полей и меньшее пространственное разрешение. Рецептивные поля соседних сенсорных нейронов могут частично перекрывать друг друга, поэтому информация о действующих на них стимулах передается не по одному, а по нескольким параллельным аксонам, что повышает надежность ее передачи.
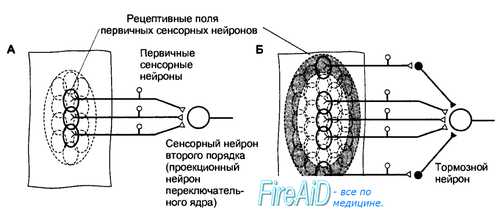 Рис. 17.1. Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Б. Раздражение центральной или периферической области рецептивного поля сенсорного нейрона второго и следующих порядков сопровождается противоположным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение). В результате контраста, создаваемого между центром и периферией рецептивного поля, выделяется информация для передачи на следующий иерархический уровень.
Рис. 17.1. Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Б. Раздражение центральной или периферической области рецептивного поля сенсорного нейрона второго и следующих порядков сопровождается противоположным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение). В результате контраста, создаваемого между центром и периферией рецептивного поля, выделяется информация для передачи на следующий иерархический уровень.
Величина рецептивных полей сенсорных нейронов второго и следующих порядков больше, чем у первичных сенсорных нейронов, поскольку центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предшествующего уровня. От центра рецептивного поля информация передается непосредственно к сенсорным нейронам следующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, поэтому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В результате сигналы от центра рецептивного поля беспрепятственно достигают следующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая функциональная организация рецептивных полей обеспечивает выделение наиболее значимых сигналов, легко различаемых на контрастном с ними фоне.
Вопрос 9. Рецептивное поле рефлекса, время рефлекса, его зависимость от силы раздражения.




![]()
![]()
Время рефлекса зависит от силы раздражения и возбудимости ЦНС.
Нервный центр. Каждый рефлекс имеет свою локализацию в ЦНС, т. е. тот ее участок, который необходим для его осуществления. При разрушении соответствующего участка рефлекс отсутствует. Однако выяснилось, что для регуляции рефлекса, его точности недостаточно первичного, или главного, центра, а необходимо участие и высших отделов ЦНС, включая кору большого мозга.

Особенности нервных центров. Характерными особенностями нервных центров, отличающими их от нервных волокон, являются быстрая утомляемость, очень высокий обмен веществ, т. е. высокая потребность в кислороде и питательных веществах, и избирательная чувствительность к некоторым ядам. Вследствие этих особенностей нарушения кровоснабжения и изменения температуры тела прежде всего сказываются на функции ЦНС.
Возбуждением называют нервный процесс, который либо вызывает деятельность органа, либо усиливает существующую.
Под торможением понимают такой нервный процесс, который ослабляет либо прекращает деятельность или препятствует ее возникновению. Взаимодействие этих двух активных процессов лежит в основе нервной деятельности.
При сокращении группы мышц тормозятся центры мышц-антагонистов. Так, при сгибании руки или ноги центры мышц-разгибателей затормаживаются. Если бы этого не произошло, то возникла бы механическая борьба мышц, судороги, а не приспособительные двигательные акты.
Понятие о доминанте. В ЦНС под влиянием тех или иных причин может возникнуть очаг повышенной возбудимости, который обладает свойством притягивать к себе возбуждения с других рефлекторных дуг и тем самым усиливать свою активность и тормозить другие нервные центры. Это явление носит название доминанты. Доминанта относится к числу основных закономерностей в деятельности ЦНС. Она может возникнуть под влиянием различных причин: голода, жажды, инстинкта самосохранения или размножения.
Важным фактором координации рефлексов является наличие в ЦНС функциональной субординации, т. е. определенного соподчинения между ее отделами, возникшего в процессе длительной эволюции. Нервные центры и рецепторы головы «авангардной» части тела, прокладывающей путь организму в окружающей среде, развиваются быстрее. Высшие отделы ЦНС приобретают способность изменять активность и направление
деятельности нижележащих отделов. Нет функций в организме, которые бы не поддавались решающему регулирующему влиянию коры больших полушарий.
По биологическому значению рефлексы подразделяются на пищевые, оборонительные, ориентировочные (ознакомление с изменяющимися условиями среды), половые (продолжение рода).
В зависимости от рабочего органа, участвующего в ответной реакции, рефлексы подразделяются на двигательные, секреторные, сосудистые.
В зависимости от продолжительности различают фазные и тонические рефлексы. Тонические рефлексы длительные, продолжаются часами, например рефлекс стояния. Любое животное может стоять часами благодаря длительному сокращению мышц. Все позные рефлексы относятся к тоническим. Они фиксируют определенное положение тела, а на их фоне разыгрываются другие, короткие, фазные рефлексы, обеспечивающие все виды рабочих, спортивных и других движений.
По принципу эффекторной иннервации рефлексы можно разделить на скелетно-моторные, или соматические (обеспечивающие двигательные акты скелетной мускулатуры), и вегетативные (функции внутренних органов).
В зависимости от того, являются ли рефлексы врожденными или приобретенными в процессе индивидуальной жизни, И. П. Павлов подразделял их на безусловные (врожденные) и условные (приобретенные).
Рецептивное поле рефлекса это
Такой метод был разработан И. П. Павловым – метод условных рефлексов, с помощью которого он расширил рефлекторную теорию, показав, что наиболее сложные и совершенные формы поведения осуществляются на основе условнорефлекторной деятельности.
Рефлексы можно классифицировать по различным показателям. По биологическому значению рефлексы подразделяются на ориентировочные, оборонительные, пищевые и половые. По расположению рецепторов они делятся на зкстерорецептивные – вызываемые раздражением рецепторов, расположенных на внешней поверхности тела; интерорецептивные – вызываемые раздражением рецепторов внутренних органов и сосудов; проприорецептивные – возникающие при раздражении рецепторов, находящихся в мышцах, сухожилиях и связках. В зависимости от органов, которые участвуют в формировании ответной реакции, рефлексы могут быть двигательными (локомоторными), секреторными, сосудистыми и др. В зависимости от того, какие отделы мозга необходимы для осуществления данного рефлекса, различают: спинальные рефлексы, для которых достаточно нейронов спинного мозга; бульбарные (возникающие при участии продолговатого мозга); мезэнцефальные (участвуют нейроны среднего мозга); диэнцефальные (нейроны – промежуточного мозга); кортикальные (для которых необходимы нейроны коры головного мозга). Следует отметить, что в большинстве рефлекторных актов участвуют как высший отдел ЦНС – кора головного мозга, так и низшие отделы одновременно.
Рефлексы можно также разделить на безусловные (врожденные) и условные (приобретенные в процессе индивидуальной жизни).
Структурной основой рефлекса, его материальным субстратом является рефлекторная дуга – нейронная цепь, по которой проходит нервный импульс от рецептора к исполнительному органу (мышце, железе).
В состав рефлекторной дуги входят:
1. воспринимающий раздражение рецептор;
2. чувствительное (афферентное) волокно (аксон чувствительного нейрона), по которому возбуждение передается в ЦНС;
3. нервный центр, в который входят один или несколько вставочных нейронов;
4. эфферентное нервное волокно (аксон эфферентного нейрона), по которому возбуждение направляется к органу.
В рефлекторной реакции всегда участвуют афферентные нейроны, передающие импульсы от рецепторов (например, проприорецепторов) исполнительного органа в ЦНС. С помощью обратной афферентации происходит коррекция ответной реакции нервными центрами, регулирующими данную функцию. Поэтому понятие «рефлекторная дуга» заменяется в настоящее время представлением о рефлекторном кольце, поскольку в функциональном отношении дуга замкнута и на периферии, и в центре беспрерывно циркулирующими во время работы органа нервными сигналами.
Простейшая рефлекторная дуга (моносинаптическая) состоит из двух нейронов: чувствительного и двигательного. Примером такого рефлекса является коленный рефлекс. Большинство рефлексов включают один или несколько последовательно связанных вставочных нейронов и называются полисинаптическими. Наиболее элементарной полисинаптической дугой является трехнейронная рефлекторная дуга, состоящая из чувствительного, вставочного и эфферентного нейронов. В осуществлении пищевых, дыхательных, сосудодвигательных рефлексов участвуют нейроны, расположенные на разных уровнях – в спинном, продолговатом, среднем и промежуточном мозге, в коре головного мозга.
Рефлексы возникают под влиянием специфических для них раздражителей, действующих на их рецептивное поле. Рецептивным полем рефлекса называется участок тела, содержащий рецепторы, раздражение которых всегда вызывает данную рефлекторную реакцию. Так, рефлекс сужения зрачка возникает при освещении сетчатки глаза, разгибание голени наступает при нанесении легкого удара по сухожилию ниже колена и т. д.
Нервным центром называется функциональное объединение нейронов, обеспечивающее осуществление какого-либо рефлекса или регуляцию какой-либо определенной функции. Нейроны, входящие в нервный центр, обычно находятся в одном отделе ЦНС, но могут располагаться и в нескольких. Центр дыхания располагается в средней трети продолговатого мозга, центр мочеиспускания – в крестцовом, центр коленного рефлекса – в поясничном отделе спинного мозга. В осуществлении сложных рефлексов целостного организма принимают участие, как правило, не один, а многие центры, расположенные в разных отделах мозга, включая его высшие отделы. Например, в акте дыхания участвует не только центр дыхания в продолговатом мозге, но и нервные клетки варолиева моста, коры головного мозга и мотонейроны спинного мозга.
Особенности распространения возбуждения в ЦНС в основном определяются свойствами нервных центров:
1. Одностороннее проведение возбуждения. В ЦНС возбуждение может распространяться только в одном направлении: от рецепторного нейрона через вставочный к эфферентному нейрону, что обусловлено наличием синапсов.
2. Более медленное проведение возбуждения по сравнению с нервными волокнами. Промежуток времени от момента нанесения раздражения на рецептор до ответной реакции исполнительного органа называется временем рефлекса. Большая его часть тратится на проведение возбуждения в нервных центрах, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5–2 мс (синоптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. Суммация возбуждений (или торможения). Нервные центры могут суммировать афферентные импульсы, что проявляется в усилении рефлекса при увеличении частоты раздражений или числа раздражаемых рецепторов. Различают два вида суммации: временная суммация – если импульсы приходят к нейрону по одному и тому же пути через один синапс с коротким интервалом, то происходит суммирование ВПСП на постсииаптической мембране и она деполяризуется до уровня, достаточного для генерации ПД; пространственная суммация связана с суммированием ВПСП, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД.
4. Конвергенция. В нервном центре несколько клеток могут передавать импульсы к одному нейрону, т. е. возбуждения конвергируют на нем. Конвергенция может быть результатом прихода возбуждающих или тормозных входных сигналов от различных источников. Так, моторные нейроны спинного мозга могут получать импульсы: от периферических нервных волокон, входящих в спинной мозг; волокон, соединяющих сегменты спинного мозга; кортикоспинальных волокон от коры мозга; тормозных путей от ретикулярной формации. В результате конвергенции происходит суммация импульсов от этих источников и возникает ответ, являющийся суммарным эффектом разнородной информации.
5. Дивергенция и иррадиация. Возбуждение даже единственного нервного волокна, по которому импульсы поступают в нервный центр, может послужить причиной возбуждения множества выходящих из центра нервных волокон. Морфологическим субстратом широкого распространения импульсов (иррадиации) возбуждения является ветвление аксонов и наличие большого числа вставочных нейронов в пределах центра.
6. Облегчение и окклюзия. На нейронах нервных центров оканчиваются не только волокна их собственных афферентных входов. Каждый из них получает веточки от афферентов соседнего центра, что может обусловливать развитие окклюзии (закупорки) или, наоборот, облегчения.
Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при раздельном раздражении каждого афферентного входа в отдельности. Явление окклюзии приводит к снижению силы суммарной ответной реакции. Феномен центрального облегчения характеризуется противоположным эффектом. На облегчении основано проторение пути – распространение возбуждения не по той цепи нейронов, по которой информация не поступала ранее, а через нейроны, уже облегченные первым раздражением.
Рецептивное поле рефлекса это




Общая физиология центральной нервной системы
Ответная реакция организма на раздражение из внешней или внутренней среды, осуществляющаяся при участии ЦНС, называется рефлексом. Путь, по которому проходит нервный импульс от рецептора до эффектора (действующий орган), называется рефлекторной дугой.
Рефлекс обеспечивает тонкое, точное и совершенное уравновешивание организма с окружающей средой, а также контроль и регуляцию функций внутри организма. В этом его биологическое значение. Рефлекс является функциональной единицей нервной деятельности.
Вся нервная деятельность складывается из рефлексов различной степени сложности, т. е. является отраженной, вызванной внешним поводом, внешним толчком. Рефлекторный принцип нервной деятельности был открыт великим французским философом, физиком и математиком Рене Декартом в XVII веке.
И. М. Сеченов писал: «. если выключить все рецепторы, то человек должен заснуть мертвым сном и никогда не проснуться.» Это теоретическое положение нашло свое обоснование в клинической практике. С. П. Боткин наблюдал больного, у которого из всех рецепторов тела функционировали один глаз и одно ухо. Как только больному закрывали глаз и затыкали ухо, он засыпал.
То же самое можно наблюдать при разрушении любого другого звена: ЦНС, чувствительных или двигательных нервных волокон. Самое сильное раздражение не вызовет ответной реакции, нервная деятельность будет отсутствовать.
Время рефлекса зависит от силы раздражения и возбудимости ЦНС. При сильном раздражении оно короче, при снижении возбудимости, вызванном, например, утомлением, время рефлекса увеличивается, при повышении возбудимости значительно уменьшается.
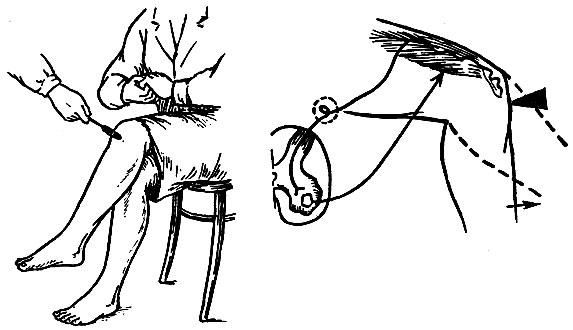
Рис. 105. Методика воспроизведения проприоцептивных рефлексов и схема рефлекторной дуги коленного рефлекса
Классификация рефлексов. Различают следующие виды рефлексов.
1. По биологическому значению рефлексы подразделяются на пищевые, оборонительные, ориентировочныеи(ознакомление с изменяющимися условиями среды), половые (продолжение рода).
3. В зависимости от рабочего органа, участвующего в ответной реакции, рефлексы подразделяются на двигательные, секреторные, сосудистые.
5. В зависимости от продолжительности различают фазные и тонические рефлексы. Тонические рефлексы длительные, продолжаются часами, например рефлекс стояния. Любое животное может стоять часами благодаря длительному сокращению мышц. Все позные рефлексы относятся к тоническим. Они фиксируют определенное положение тела, а на их фоне разыгрываются другие, короткие, фазные рефлексы, обеспечивающие все виды рабочих, спортивных и других движений.
7. По принципу эффекторной иннервации рефлексы можно разделить на скелетно-моторные, или соматические (обеспечивающие двигательные акты скелетной мускулатуры), и вегетативные (функции внутренних органов).
8. В зависимости от того, являются ли рефлексы врожденными или приобретенными в процессе индивидуальной жизни, И. П. Павлов подразделял их на безусловные (врожденные) и условные (приобретенные).
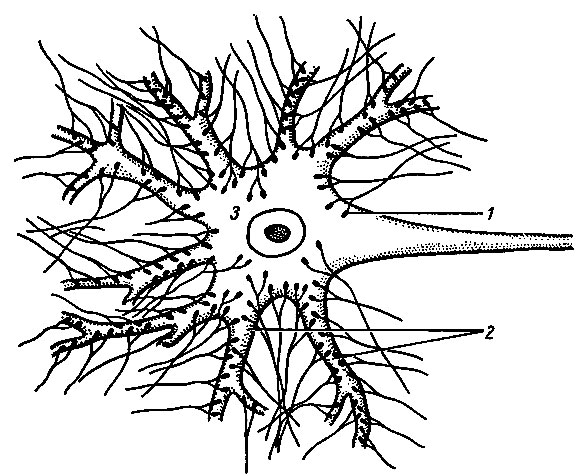
Рис. 106. Синаптические бляшки (1) окончаний пресинаптических аксонов образуют соединения на дендритах (2) и теле (3) нейрона [Стерки П., 1984]
Синапсы по механизму передачи возбуждения разделяются на химические и электрические. Последние находятся в сердечной мышце, гладких мышцах и железистой ткани; в ЦНС наличие их только предполагается.
Синапс, с химической передачей, состоит из синаптической бляшки, пресинаптической мембраны, синаптической щели шириной 30 нм и постсинаптической мембраны (рис. 107).


