Роль нейропептида Y в патогенезе сердечно-сосудистых заболеваний

Нейропептид Y (NPY) представляет собой высококонсервативный пептид, который широко распространен в центральной и периферической нервной системе. NPY в основном обнаруживается в постганглионарных симпатических нейронах, из которых он высвобождается одновременно с норэпинефрином (NE) в ответ на симпатическую стимуляцию.
NPY модулирует свои функции в сердечно-сосудистой, центральной и периферической системах посредством активации рецепторов, связанных с G-белком, связанными с Y (GPCR; Y1R, Y2R, Y3R, Y4R, Y5R и γ6R). Подобно пептиду YY (PYY) и полипептиду поджелудочной железы (PP), NPY является одним из «полипептидных полипептидов PP». Несмотря на структурное сходство, каждый «полипептидный полипептид» реализует специфические функциональные эффекты.
NPY играет особую роль в различных физиологических функциях, включая регуляцию настроения, сердечно-сосудистый и иммунный гомеостаз, вазомоцию, ангиогенез, ремоделирование сердца, аппетит, моторику желудочно-кишечного тракта, нейроэндокринную ось, симпатическую и вагусную иннервацию. NPY является наиболее распространенным нейропептидом в сердце и присутствует в постганглионарных симпатических нейронах, снабжающих сосудистую сеть, эндокард и кардиомиоциты, а также во внутрисердечных ганглиях и парасимпатических нейронах. Тем не менее, в дополнение к его важной роли в нормальных физиологических механизмах контроля, он также участвует в патогенезе ряда патологических процессов при сердечно-сосудистых заболеваний.
Нейропептид Y1 рецептор
Нейропептид Y2 рецептор
Нейропептид Y5 рецептор
NPY проявляет двойной противоположный эффект в сердечно-сосудистых тканях, включая действие в качестве кардиодепрессанта и сердечного стрессора. Эти противоречивые эффекты NPY в значительной степени зависят от характера экспрессии рецептора NPY, концентрации лиганда, активности DPP-4 и адренергической активности. Связанные с сердцем рецепторы NPY, Y1R, Y2R и Y5R, как правило, участвуют в патологических состояниях и предположительно играют важную роль в патогенезе сердечно-сосудистых заболеваний, включая гипертонию, атеросклероз, ишемию / инфаркт миокарда, диабетическую, стрессовую и гипертрофическую кардиомиопатию, и сердечная недостаточность.
Высоко экспрессируемый в симпатических нервных окончаниях вокруг сосудистой сети, NPY действует как мощный вазоконстриктор наряду с NE. Увеличение циркулирующего NPY опосредуется периферическими нервами и мозговым веществом надпочечников. Совсем недавно повышенные уровни NPY наблюдались при гипертонической беременности (преэклампсия) у женщин.
NPY (Нейропептид Y)
 Нейропептид Y (NPY) представляет собой 36-аминокислотный нейропептид, который действует как нейромедиатор в мозге и вегетативной нервной системе человека; небольшие вариации пептида можно найти и у многих других животных. В автономной системе он в основном производится нейронами симпатической нервной системы и служит в качестве сильного сосудосуживающего средства, а также вызывает рост жировой ткани. Данный нейропептид производится в различных участках мозга, включая гипоталамус, и, как полагают, имеет несколько функций, в том числе: увеличение аппетита и количества принимаемой пищи, а также сохранение энергии в виде жира, снижение тревожности и напряжения, ослабление восприятия боли. Вещество также оказывает влияние на циркадный ритм, снижает добровольное потребление алкоголя, уменьшает артериальное давление и помогает контролировать эпилептические припадки.
Нейропептид Y (NPY) представляет собой 36-аминокислотный нейропептид, который действует как нейромедиатор в мозге и вегетативной нервной системе человека; небольшие вариации пептида можно найти и у многих других животных. В автономной системе он в основном производится нейронами симпатической нервной системы и служит в качестве сильного сосудосуживающего средства, а также вызывает рост жировой ткани. Данный нейропептид производится в различных участках мозга, включая гипоталамус, и, как полагают, имеет несколько функций, в том числе: увеличение аппетита и количества принимаемой пищи, а также сохранение энергии в виде жира, снижение тревожности и напряжения, ослабление восприятия боли. Вещество также оказывает влияние на циркадный ритм, снижает добровольное потребление алкоголя, уменьшает артериальное давление и помогает контролировать эпилептические припадки.
Открытие
После изоляции нейропептида-Y (NPY) из гипоталамуса свиньи в 1982 году исследователи начали высказывать гипотезы о вовлечении NPY в гипоталамо-опосредованные функции. В исследовании 1983 года, NPY-аксонные окончания были обнаружены в паравентрикулярном ядре (PVN) гипоталамуса, а наиболее высокие уровни иммунореактивности NPY были найдены в паравентикулярном ядре гипоталамуса. Шесть лет спустя, в 1989 году, Моррис и др. сфокусировали внимание на расположении ядер NPY в мозге. Кроме того, результаты гибридизации показали, что наиболее высокие клеточные уровни NPY мРНК находятся в дугообразном ядре гипоталамуса. В 1989 году Хаас и Джордж сообщили о том, что местные инъекции нейропептида Y в паравентикулярное ядро гипоталамуса приводят к быстрому высвобождению кортикотропин-рилизинг-гормона (КРГ) в мозге крысы, что доказывает тот факт, что активность NPY непосредственно стимулирует высвобождение и синтез КРГ. Последние данные представляют собой важнейший материал в исследованиях NPY. Значительный объем работ по КРГ и его роли в стрессе и расстройствах пищевого поведения, таких как ожирение, был проведен в 1970-х гг. Эти исследования, в совокупности, положили начало пониманию роли NPY в орексигенезе или приеме пищи.
Роль NPY в приеме пищи
Для подтверждения того факта, что повышение активности NPY действительно увеличивает потребление пищи, были проведены поведенческие анализы в исследованиях пищевого поведения крыс, совместно с иммунологическими и полевыми исследованиями гибридизации. Исследования показывают, что стимуляция активности NPY через введение некоторых агонистов NPY увеличивает потребление пищи у крыс по сравнению с исходными данными. Воздействие активности NPY на потребление пищи также демонстрируется блокадой определенных рецепторов нейропептида Y (рецепторов Y1 и Y5), которые, как ожидалось, ингибируют активность NPY, уменьшая, таким образом, потребление пищи. Тем не менее, исследование 1999 года, проведенное Кингом и соавторами, продемонстрировало эффекты активации NPY ауторецептора Y2, который, как было показано, ингибирует высвобождение NPY и таким образом регулирует потребление пищи после его активации. В этом исследовании высокоселективный антагонист Y2, BIIE0246, вводили местно в ядро гипоталамуса. Радиоиммунологические данные после инъекции BIIE0246 показывают значительное увеличение высвобождения NPY по сравнению с контрольной группой. Хотя фармакологический полураспад экзогенного NPY, других агонистов и антагонистов по-прежнему остается латентным, этот эффект не является длительным и организм крысы способен регулировать и нормализовать аномальные уровни NPY и, следовательно, потребление пищи.
Роль NPY в ожирении
В1995 году Драйден и соавт. провели исследование с использованием генетически тучных крыс, чтобы продемонстрировать роль NPY в расстройствах пищевого поведения, таких как ожирение. Исследование выявило четыре основных фактора, способствующих ожирению у крыс:
Кроме того, эти факторы также коррелируют друг с другом. Устойчивые повышенные уровни глюкокортикостероидов стимулируют глюконеогенез, что впоследствии приводит к повышению уровня глюкозы в крови, активируя высвобождение инсулина для регулирования уровня глюкозы, вызывая его обратный захват и сохраняя в виде гликогена в тканях организм. В случае ожирения, которое, как полагают исследователи, имеет мощные генетические и диетические предпосылки, резистентность к инсулину снижает регулирование глюкозы в крови, что приводит к изменениям уровней глюкозы и развитию сахарного диабета. Кроме того, высокие уровни глюкокортикоидов вызывают увеличение NPY путем прямой активации глюкокортикостероидных рецепторов II типа (которые активизируются только при относительно высоких уровнях глюкокортикостероидов) и, косвенно, путем отмены отрицательной обратной связи кортикотропин-рилизинг фактора (КРФ) при синтезе и высвобождении NPY. Между тем, индуцированная ожирением резистентность к инсулину и мутация рецептора лептина приводит к отмене ингибирования активности NPY и, в конечном счете, потребления пищи, через другие негативные механизмы обратной связи для их регулирования. При адреналэктомии или гипофизэктомии ожирение у крыс значительно сокращалось.
Корреляция со стрессом и диетой
Исследования на мышах и обезьянах показали, что повторяющиеся стрессы и диета с высоким содержанием жира и сахара стимулируют высвобождение нейропептида Y, в результате чего наблюдается скопление жира на животе. Исследователи полагают, что, управляя уровнями NPY, можно убирать жир из областей, где он нежелателен и перемещать его туда, где он необходим. Более высокие уровни NPY могут быть связаны с устойчивостью к и восстановлением от посттравматического стрессового расстройства. При ослаблении реакций страха они позволяют людям лучше работать в ситуациях крайнего напряжения.
NPY и алкоголизм
Результаты показывают, что NPY может выступать в качестве средства от алкоголизма:
Рецептор нейропептида Y
Рецепторный белок, с которым работает NPY, является G-протеин связанным рецептором из родопсиноподобной 7-трансмембранной GPCR семьи. У млекопитающих было выявлено пять подтипов рецепторов NPY, четыре из которых функционируют в организме человека. Подтипы Y1 и Y5 играют важную роль в стимулировании вскармливания, в то время как Y2 и Y4 важны при ослаблении аппетита (обеспечении чувства сытости). Некоторые из этих рецепторов являются одними из наиболее сохраняемых нейропептидных рецепторов.
Нейропептиды
Нейропептиды (эндогенные опиоиды, энкефалины) представляют собой так называемую третью группу высокоактивных медиаторов. Они широко представлены в ЦНС, особенно в задних рогах спинного мозга, базальных ганглиях и вегетативной нервной системе. Синтез и высвобождение нейропептидов происходит в разных участках нейрона.
Пептиды синтезируются в рибосомах, расположенных в теле нейрона, а не в аксонах-нервных терминалях. После синтеза они упаковываются в крупные плотные пузырьки в аппарате Гольджи, затем медленно переносятся в терминаль с помощью аксонного транспорта, также медленно они оставляют синапс.
Гонгломераты пептидов в виде крупных белков предшественников, часто содержат последовательности нескольких биологически активных пептидов (Mains R., Eipper B., 1999). Низкомолекулярные медиаторы (ацетилхолин, дофамин и др.), образующиеся внутри аксонных окончаний, напротив, синтезируются быстро, а их количество в синапсе оперативно контролируется структурами нервного терминаля.
 | Для выявления влияния нейромедиаторов на развитие шизофрении необходимо проводить лабораторные обследования пациентов |
Количество пептида, доступное для выделения, ограничено его количеством в терминале. Однако связывание пептидов с рецепторами происходит при более низких концентрациях, чем связывание низкомолекулярных медиаторов, таких, например, как ацетилхолин. Рецепторы нейропептидов, как и другие метаботропные рецепторы, действуют через внутриклеточные пути, обеспечивающие значительное усиление сигналов, поэтому для достижения эффекта на постсинаптические рецепторы требуется небольшое количество молекул пептида. Потребность в необходимом для высвобождения количестве нейропептида может быть обеспечена транспортом молекул из тела клетки.
Нейропептиды могут модулировать действие классических медиаторов. Так, в частности, действие катехоламинов часто модулируется нейропептидами — эндогенными опиоидами эндорфинов, которые способны изменять дофаминергическую трансмиссию.
Нейропептиды
Поскольку изменение пептидергических систем отмечено при шизофрении, была высказана гипотеза, согласно которой первичным при шизофрении следует считать нарушение функционирования эндорфинов (синтеза\распада), которые в дальнейшем опосредованно влияют на дофаминергические синапсы, приводя к торможению дофаминергической передачи.
Дофаминергические окончания включают в себя и другие нейропептиды, обладающие модулирующим эффектом. Это пептиды, тормозящие дофаминергическую передачу, — холицистокинин-8 и (в меньшей степени) соматостатин. Возможностью активировать трансмиссию обладает вазоинтенстинальный пептид, в то время как нейропептид Y может активировать норадренергическую передачу.
В гиппокампе мозга больных шизофренией обнаружено снижение концентрации холицистокинина-8 и соматостатина, а в миндалине уменьшено содержание холисцистокинина-8, мет — энкефалина и вещества Р, повышен уровень нейропептида Y.
Исследования показали, что эндорфины и дезтирозил — эндорфины, холицистокинин-8, близкий к последнему пептид церулин обладают нейролептическим эффектом. Эти соединения могут быть использованы в комплексном лечении больных шизофренией.
Перспективны попытки использования для терапии шизофрении синтетических антагонистов производного холицистокинина-8-С-концевого тетрапептида (холицистокинин-4), влияющих на те дофаминергические нейроны, от активности которых зависит состояние тревоги.
Не вызывает сомнения, что в генезе шизофрении принимают участие различные нейротрансмиттеры. В настоящее время большинство ученых, хотя и медленно, отходят от исключительной концентрации внимания лишь на нарушении активности дофаминергических нейронов при этом заболевании. Изучаются искаженные взаимодействия между системами нейромедиаторов, при этом в центре внимания по-прежнему остаются дофамин, серотонин и глутамат.
PsyAndNeuro.ru
Таинственный мир нейропептидов

Гормоны являются самыми известными регуляторами процессов в организме. Их функции и химическая структура изучены достаточно хорошо, что позволяет создавать на их основе лекарственные препараты и успешно проводить, например, заместительную терапию. На втором месте по освещенности располагаются нейромедиаторы. Эти соединения преимущественно осуществляют связь нервных клеток друг с другом, таким образом активируя или угнетая различные области и процессы. Влиять на нейромедиаторы несколько сложнее, однако разработано множество препаратов, воздействующих на метаболизм, выброс, обратный захват и т.д., следовательно, нейромедиаторная система поддается регулировке.
На данном этапе развития медицины и фармакологии менее всех изучены и подвергаются контролю нейропептиды. Тем не менее они принимают активное участие практически во всех процессах в организме, регулируют самые важные поведенческие и приспособительные реакции, управляют циркадными ритмами, пищевым и половым поведением, воздействуют на гормоны и нейромедиаторы. Обойти вниманием данную тему невозможно, как и собрать воедино все данные по ней. По результатам проводимых исследований данные не всегда совпадают, иногда же получаются прямо противоположными, что значительно затрудняет описание нейропептидов, за счет отсутствия единой классификации возникает еще большая путаница. Таким образом сведения о нейропептидах недостаточно систематизированы, тем не менее данный материал является наиболее полным на русском языке и включающим в себя значительное количество соединений, не указанных в других обзорах.
Нейропептиды (НП) – биологически активные соединения, которые участвуют в регуляции обмена веществ, поддержании гомеостаза, воздействуют на иммунные процессы, играют важную роль в механизмах памяти, обучения, сна и т.д. Для НП характерна относительно короткая последовательность цепи, состоящей из аминокислотных остатков, как правило, она составляет 5-52 элементов [19].
НП образуются в результате последовательного гидролитического разложения пептида-предшественника, так, из одной исходной молекулы может образоваться несколько веществ, которые на каждом из этапов могут обладать биологической активностью, отличающейся от начального и конечного продуктов. Предшественники НП синтезируются в теле клетки путем трансляции гена, кодирующего пептид, далее протеазы точечно расщепляют длинную молекулу на более короткие цепи, которые в последующем могут подвергнуться трансформации или остаться в неизмененном виде. Если речь идет о нейроне, то далее НП транспортируется в пресинаптическую терминаль, откуда выделяется в синаптическую щель. Некоторые нейропептиды способны выполнять функции медиаторов, осуществляя непосредственную передачу нервного импульса, другие изменяют метаболизм клетки, выполняя роль нейромодулятора. НП способны передавать сигнал не только на близкие расстояния, рецепторы к некоторым из них находятся на достаточном удалении от места синтеза, что позволяет сравнивать их с гормонами. Функции многих НП дублируют друг друга, тем не менее каждый имеет уникальный спектр активности. Эта особенность позволяет более точно передавать сигнал и регулировать взаимодействие различных частей организма, координировать сложную систему непрерывно происходящих процессов, так или иначе влияющих друг на друга и требующих постоянных коррекций. Подавляющее большинство нейропептидов воздействует на «медленные» метаботропные рецепторы, связанные с G-белками. Кроме того, НП способны изменять активность друг друга и некоторых гормонов (чаще ингибировать или активировать их синтез), что приводит к запуску каскадных реакций [10, 12].
Многие нейропептиды синтезируется в большей или меньшей степени в различных органах и тканях, и только некоторые из них строго специфичны для определенных зон. Распространение НП в организме неоднородно, существуютместа, наиболее характерные для каждого вещества, тем не менее в незначительных количествах они определяются практически во всех тканях. В нервной ткани НП присутствуют в немиелинизированных волокнах С-типа и небольших миелинизированных волокнах А-дельта-типа. В спинном мозге НП синтезируются клетками дорсальных рогов ганглиев, далее транспортируются по аксонам в нервные окончания, где могут выступать в роли нейромедиаторов. В синаптических терминалях НП способны оказывать действие совместно с нейромедиаторами небелковой природы. Нейропептид может быть колокализован с одним или несколькими медиаторами, что приводит к некоторому усилению или изменению действия. Если их выделение совпадает по времени, то эффект зависит от них обоих, но также они способны выделяться раздельно, что приводит к реализации биологического эффекта каждого из них отдельно друг от друга [16].
Полной и учитывающей все многообразие НП классификации на данный момент не существует. Делались попытки разработать ее на основании химической структуры, функций или места синтеза. Однако многие НП способны выполнять несколько функций в зависимости от места нахождения, сходные по структуре соединения – отвечать за разные процессы, а различные вещества по происхождению быть агонистами. НП найденные в каких-либо тканях, не всегда были для них специфичны и в дальнейшем были обнаружены в других органах. Кроме того, на настоящий момент открываются новые соединения, которые невозможно отнести к какой-либо из существующих групп, так как не до конца изучены их функции. Наиболее полной и функциональной можно считать классификацию НП по семействам, так как она учитывает наибольшее количество особенностей этих веществ [2, 8].
Классификация НП
Гипоталамические либерины и статины
Тиролиберин, кортиколиберин, люлиберин, соматолиберин, соматостатин, меланостатин.К первому семейству относятся нейропептиды, синтезируемые гипоталамусом. Они были объединены в одну группу по топологическому признаку и имели преимущественно функцию стимуляции (либерины) или угнетения (статины) синтеза гормонов гипофиза. По химической структуре гипоталамические нейропептиды значительно друг от друга отличаются и имеют различных предшественников. Кроме воздействия на удаленные от места синтеза структуры, НП данной группы могут влиять на соседние нейроны, угнетая или наоборот стимулируя образование друг друга. По мере изучения, представители первого семейства были найдены в других органах и тканях и обнаружили способность влиять на эмоциональное состояние, пищевое и половое поведение, регуляцию цикла сон-бодрствование, обеспечение и запуск стресс-протективных механизмов, стимуляцию иммунных процессов, нейрогенеза и многие другие [10].
Опиоидные пептиды
Семейство опиоидных пептидовимеет характерную для большинства представителей последовательность аминокислот Tyr-Gly-Gly-Phe. Соответственно ОП воздействует на опиоидные рецепторы преимущественно μ (MOP), δ (DOP), и κ (KOP), ассоциированные с G-белками. За счет способности связываться с перечисленными рецепторами, ОП обладают налоксон-ингибирующим, морфиноподобным анальгезирующим и седативным действием.ОП обладают разнообразными биологическими эффектами. В отношении воздействия на поведение, у них выделяют способность воздействовать на агрессию, мотивацию удовлетворения, половое влечение, пищевое насыщение, стрессорные адаптивные процессы, лекарственную зависимость, седацию, модуляцию болевой чувствительности и т.д. Кроме того они участвуют в нейродегенеративных процессах, повреждении ткани мозга вследствие травмы и ишемии [6, 8, 10, 13, 16].
Опиоидные пептиды (ОП) широко распространены в центральной и периферической нервной системах, ЖКТ, сыворотке крови, вырабатываются не только нейронами, но и клетками эндокринной и иммунной систем. Большая часть опиоидных пептидов образуется из общих белковых предшественников – проопиомеланокортин (α-MSH, γ-MSH, β-MSH, АКТГ, β-эндорфин, α-эндорфин, γ-эндорфин, β-липопротеин (β-LPH), γ-LPH, CLIP), продинорфин (динорфин А, динорфин В, α-неоэндорфин, β-неоэндорфин, динорфин-32, лейморфин), препроэнкефалин (лей-энкефалин, мет-энкефалин, амидорфин, адренорфин, пептидВ, пептидЕ, пептидF), препроноцицептин, препроорфарин (ноцицептин (орфанин FQ)), препро-NPFF (НП FF, НП AF, НП SF) и др.[20].

Энкефалины представляют собой короткие пептидные цепи, от 5 аминокислотных остатков. Типичные члены семейства – лей-энкефалин и мет-энкефалин, названные по пятой аминокислоте, соответственно лейцин и метионин.Кроме них в состав группы входят DTLET и DAMGO. Воздействуют преимущественно на δ-опиоидные рецепторы. Оба нейропептида обладают выраженным морфиноподобным анальгезирующим, седативным действием. Принимают участие в формировании поведенческих реакций. Доказано их участие во многих нейродегенеративных патологиях.
Эндорфины α и β являются продуктами гидролиза ПОМК и содержат 16 и 31 аминокислотных остатков соответственно. Участвуют в мотивации алкогольного поведения, ноцицептивных реакциях, стрессовом ответе, регуляции циркадных ритмов. Β-эндорфин является менее специфичным по отношению к рецепторам и может активировать в большей или меньшей степени все три из числа опиоидных.
Динорфины образуются в процессе преобразования продинорфина и содержат в своей структуре последовательность лей-энкефалина. Наиболее важным действием является центральный и периферический ноцицептивный процесс. В состав группы входят динорфин А и динорфин В (риморфин), содержащие соответственно 17 и 13 аминокислотных остатков. Кроме них образуются α-, β-неоэндорфин. Все они активируют преимущественно κ-опиоидные рецепторы. Их селективность обусловлена наличием аргинина и лизина на С-конце. Если протеолиз предшественника происходит не полностью, то образуется так называемый «большой» динорфин, в состав которого входят динорфин А и В, и обладающий такими же свойствами, но отличающийся большей селективностью в отношении КОР [15].
Дерморфин и дельторфинспецифические агонисты µ- и δ- опиоидных рецепторов соответственно, состоящие из 7 аминокислотных остатков. Участвуют в снижении порога эпилептической готовности, оказывают выраженныйанальгезирующий эффект, стимулируют выброс β-эндорфина. Отличаются присутствием D-аминокислоты во втором положении, что обусловливает их большую устойчивость к ферментативному гидролизу.
Геморфины являются продуктами протеолитического распада гемоглобина, обладают сродством к µ-опиоидным рецепторам. Участвуют в анальгезирующей реакции и развитии эйфории после физической нагрузки.
Β-казаморфин состоит из 7 аминокислотных остатков и образуется путем гидролиза казеина. Способен активировать µ-опиоидные рецепторы, вызывать стимуляцию иммунной системы и повышать потребление пищи.
Леуморфин (лейморфин) образуется из предшественника препродинорфина. Обладают достаточным сродством к опиоидным рецепторам и по биологическому действию сопоставим с другими ОП.
Адренорфинсинтезируется преимущественно в надпочечниках. Обладает способностью влиять на ноцицептивные процессы, как и другие пептиды семейства.
Нейропептиды AF и SF состоят из 18 и 11 аминокислотных остатков соответственно.НейропептидFF состоит из 8 аминокислотных остатков. Рецепторы к нему расположены в основном в спинальной и супраспинальной зонах, содержащих большое количество эндогенных опиоидов.
Ноцицептин (орфанин FQ) – состоит из 17 аминокислотных остатков и имеет структуру аналогичную с опиоидными пептидами. Рецепторы к ноцицептину похожи на опиоидные, связаны с аденилатциклазой. При воздействии на данные рецепторы происходит активация калиевых каналов и ингибирование кальциевых. Ноцицептин и рецепторы к нему наиболее широко представлены в коре, ольфакторных ядрах, амигдале, гиппокампальной формации и дорзальных рогах спинного мозга. Принимает участие в процессах памяти, обучения, стрессовых реакциях. В экспериментальных моделях показал способность снижать уровень тревоги. Активация рецепторов ноцициептина приводит к анальгезии, но препятствует действию опиоидов[13].
Меланокортины
Адренокортикотропин (АКТГ) – гормон, синтезирующийся в передней доле гипофиза, основной функцией которого является стимуляция продукции надпочечниками кортикостероидов. Доказано, что он способен синтезироваться и другими отделами мозга, и кроме гормональной функции выступать в роли нейромедиатора, принимать участие в регуляции высших корковых функций, таких как память, внимание, обучение.
А- β- γ-меланотропины (меланоцитостимулирующие гормоны)образуются из проопиомеланокортина. Синтез происходит наиболее интенсивно в средней доле гипофиза. Рецепторы к МСГ ассоциированы с G-белками и разделяются на 2 типа: МСНR1 иMCHR2.Экспрессия рецепторов 1 типа наиболее высока в коре, гиппокампе, миндалине и прилежащем ядре, что позволяет предположить, что данные нейропептиды принимают участие в развитии таких патологий как расстройства настроения и шизофрения. Это подтверждается введением антагонистов к этому типу рецепторов, что вызывало анксиолитический и антидепрессивный эффекты. Α-МСГ стимулирует образование пигмента в коже, участвует в психических процессах – памяти и обучении, сна, агрессии, модулирует воспаление в мозге, блокирует синтез глией фактора некроза опухоли. Γ-МСГв меньшей степени влияет на пигментный обмен, но усиливает стероидогенную функцию АКТГ. Все МСГ способны участвовать в регуляции функций ЖКТ, иммунных процессов, клеточного роста и митоза, пищевого поведения [2, 16, 19].
Вазопрессин-тоцины
Вазопрессин и окситоцин вырабатываются в гипоталамусе, по аксонам попадают в заднюю долю гипофиза, откуда выделяются в кровь. Оказывают сильное действие на формирование поведенческих реакций, таких как привязанности, половое, родительское поведение. При стрессе участвуют в создании защитных механизмов. Кроме того, способны влиять на артериальное давление, сокращение гладкой мускулатуры, обмен веществ.
Мезотоцин, изотоцин, вазотоцинсостоят из 10 аминокислотных остатков, имея общую начальную последовательность из 6 аминокислот. Синтезируются преимущественно в задней доле гипофиза вместе с окситоцином и вазопрессином и похожи на них по биологическим эффектам, однако менее активны [2, 8, 10].
Предшественники – препровазопрессин-нейрофизин II (вазопрессин, нейрофизин II), препроокситоцин-нейрофизин I (окситоцин, нейрофизин I) [20].
Панкреатические пептиды
Нейропептид Yсостоит из 36 аминокислотных остатков. Распространен в мозге (гипоталамической и кортикальной областях, гиппокампе, таламусе) и периферической нервной системе, постганглионарных симпатических волокнах, надпочечниках, мегакариоцитах и тромбоцитах. Имеются данные о изменении распределения НП в популяции нейронов префронтальной коры в онтогенезе при патологических нарушениях. Угнетает выделение трансмиттера из нервных окончаний. Действие проявляется гипотензией, гипотермией и угнетением дыхания, секреции воды и электролитов в кишечнике, регуляцией циркадных ритмов и мотивационного поведения. Способен участвовать в регуляции пищевого поведения, так, при его хроническом введении в ЦНС масса тела увеличивалась. В период уменьшения потребления пищи, количество нейропептида Y в дугообразном и паравентрикулярных ядрах увеличивалось. При распаде нейропептида Y, получившиеся вещества могут выступать как его агонисты и антагонисты, что определяется их химической структурой.
Пептид тирозин-тирозин (PYY) имеет сходную структуру с нейропептидом Y и отличается от него дополнительным аминокислотным остатком –Tyr. По выполняемым функциям они так же похожи и конкурируют за связывание с одними и теми же рецепторами.
Панкреатический полипептид (РРY) синтезируется РР-клетками островков Лангерганса поджелудочной железы. Имеет в своем составе 36 аминокислотных остатков. По механизму действия является антагонистом холецистокинина, подавляет секреторную активность клеток поджелудочной железы и стимулирует выработку желудочного сока, задерживая пищу в желудке [8, 14, 16].
Предшественники – препро-NPY(NPY), препро-PPY(PPY), препро-PYY(PYY) [20].
Глюкагон-секретины
Глицентинявляется промежуточным продуктом образования глюкагона из препроглюкагона. До конца его свойства не изучены, предполагается, что спектр его активности похож на таковой глюкагона.
Глюкагон (GRP) содержит 27 аминокислотных остатков. В наибольших количествах содержится в ЖКТ и мозге, секретируется некоторыми опухолями, где стимулирует митоз, оказывает трофическое влияние на нормальные и неопластические клетки, включая аутокринную стимуляцию пролиферации клеток мелкоклеточного рака легких. Участвует в регуляции обмена глюкозы, при повышении ее уровня стимулирует синтез инсулина и тормозит пищевое поведение. В ЦНС улучшает процессы формирования памяти, принимает участие в реакции организма на стресс.Участвует в регуляции циклов сон-бодрствование, температуры тела, аппетита и чувства насыщения. Модулирует активность макрофагов. Регулирует секрецию ферментов поджелудочной железы и соляной кислоты в желудке, стимулирует сокращение гладкой мускулатуры и высвобождение некоторых гормонов кишечника, включая гастрин.
VIP (вазоактивный интестинальный пептид) состоит из 28 аминокислотных остатков. Широко представлен в ЦНС, особенно в коре головного мозга, где участвует в формировании поведенческих реакций, оказывает положительное действие на процессы обучения и памяти, полового поведения. Может выступать как медиатор в серотонинергических и холинергических системах. На периферии вызывает расширение бронхов, сосудов(в том числе церебральных).
Секретинсодержит 27 аминокислотных остатков. Продуцируется преимущественно мозговыми структурами, надпочечниками и кишечником. Наиболее сильно его воздействие проявляется на клетки ЖКТ, где он приводит к расслаблению гладкой мускулатуры и повышению секреции гормонов поджелудочной железой.
Гастринпреимущественно вырабатывается G-клетками желудка и поджелудочной железой. Выделяют «большой» гастрин, который состоит из 34 аминокислотных остатков, гастрин-17 и гастрин 14, которые содержат соответственно 17 и 14 аминокислот. По выполняемым функциям они аналогичны, так как все содержат одинаковый активный центр.Увеличивают секрецию соляной кислоты, пепсина, бикарбонатов, секретина, холецистокинина, соматостатина и некоторых других пептидов, участвующих в пищеварении. Тормозит опорожнение желудка. Приводит к расширению сосудов желудка посредством увеличения продукции простагландина Е. Участвует в регуляции пищевого поведения, снижая мотивацию поиска пищи [1, 8, 10, 14, 16].
Предшественники – препрогастрин (гастрин), препроглюкагон (глицентин, глюкагон), препросекретин (секретин), препро-VIP (VIP).
Холецистокинин
Тахикинины
К данной группе относятся НП, имеющие предшественника β-препротахикинина и содержащие на С-конце последовательность-Gly-Leu-Met. Биологические эффекты опосредованы воздействием на рецепторы, ассоциированные с G-белком. Тахикинины могут выступать также в роли нейротрансмиттеров и широко представлены в различных тканях организма. Главные физиологические эффекты заключаются в регуляции тонуса гладкой мускулатуры кишечника, бронхов, принимают участие в поведенческих реакциях, ноцицептивных процессах, воспалительных процессах.
Вещество Рсодержит в свое составе 11 аминокислотных остатков. Открыт в 1931г. и является наиболее изученным из семейства. Синтезируется преимущественно в ЦНС – миндалине, перегородке, гиппокампе, гипоталамусе и сером веществе у водопровода мозга, которые участвуют в формировании тревоги и депрессии. Встречается в задних рогах спинного мозгаи является нейромодулятором в первичных афферентных волокнах и немиелинизированных волокнах С-типа. Обладает широким спектром физиологических эффектов – способен влиять на уровень артериального давления, капиллярную проницаемость, сокращение гладкой мускулатуры, обладает секретогенным действием, участвует в контроле секреции пролактина и пищеварительных гормонов. Синтез вещества Р усиливается допамином: так, выявлено, что при повреждении допаниэргических волокон, уменьшается экспрессия мРНК, отвечающая не только за образование вещества Р, но и энкефалинов и динорфина. Участвует в передаче болевого сигнала.Изучается способность вещества Р влиять на обучение, сон, устойчивость к стрессу.
Нейрокинины (А, В и К) по своим эффектам сходны с веществом Р, но имеют отличную от него специфичность к рецепторам. Изменяют возбудимость нервных клеток, способны оказывать противовоспалительный эффект – их воздействие приводит к расширению сосудов и увеличению их проницаемости, выделению тучными клетками и лейкоцитами простагландина Е2, цитокинов и аминов.Кроме указанных физиологических эффектов участвуют в передаче нервного импульса.
Кассининсостоит из 12 аминокислотных остатков. Принимает участие в снижении артериального давления, стимуляции сокращения гладкой мускулатуры. Имеются данные о том, что кассинин обладает противосудорожной активностью [1, 8, 16, 19, 20].
Мотилин
В своем составе содержит 22 аминокислотных остатка. Вырабатывается преимущественно в ЖКТ, где влияет главным образом на моторику – усиливает тонус нижнего сфинктерапищевода, стимулирует опорожнение желудка и моторику толстого кишечника. Стимулирует выработку инсулина и соматостатина поджелудочной железой. В ЦНС наибольшая концентрация обнаруживается в гиппокампе. Действие мотилина на психические функции изучено недостаточно, предполагается, что он влияет на пищевое поведение [2].
Предшественник – препромотилин [20].
Нейротензины
Нейротензин обнаруживается преимущественно в гипоталамусе, мезокортиколимбической и нигростриатной зонах, вентральной покрышке, перегородке, поясной извилине, слизистой тонкого кишечника. Пептид оказывает сильное гипотензивное действие, приводит к сокращению гладкой мускулатуры, снижает температуру тела, увеличивает содержание глюкозы и глюкагона, может связываться с рецепторами тучных клеток. Нейротензин обладает в некотором смысле гормональным действием – в гипофизе усиливает секрецию ЛГ и ФСГ. Имеются данные о его способности влиять на половое поведение, развитие стресс-реакции и ноцицептивные процессы.Установлено, что дисфункция в системе нейротензина встречается при психических заболеваниях, в частности шизофрении. Предполагается, что воздействие на его обмен может иметь эффект при психотических состояниях, не вызывая при этом повышения веса и каталептических проявлений. Однако клинически значимых подтверждений этому получено не было. Ассоциирован с дофаминергической, серотонинергической, ГАБАергической, глутаматергической и холинергической системами.
Нейромедины Nи U(NMN,NMU) секретируется в клетках ЦНС и ЖКТ. Рецепторы к ним расположены в нейронах, клетках тонкого кишечника, поджелудочной железы, желудка, лимфоцитах, моноцитах, мышечных волокнах матки. Воздействие NMU приводит к снижению массы тела посредством снижения аппетита. В регуляции иммунных процессов усиливает воспаление, активируя тучные клетки. NMN может выступать в роли нейромедиатора.
Ксенопсинучаствует в регуляции тонуса гладкой мускулатуры [8, 14, 16, 19].
Предшественник – пропренейротензин [20].
Бомбезины
Бомбезин состоит из 14 аминокислотных остатков. Является мощным активатором гипоталамо-гипофизарно-надпочечниковой оси, участвует в регуляции стресс-реакции организма, влияет на процессы запоминания. Кроме того, регулирует потребление этанола. Физиологические эффекты заключаются в вазоконстрикции, снижении температуры тела, регуляции секреторных процессов в ЖКТ, принимает участие в аутокринной стимуляции пролиферации клеток и роста малых раковых клеток легких.
Гастрин-рилизинг пептид (GRP) состоит из 27 аминокислотных остатков. Распространен как в мозговой ткани, кишечнике, легких, иммунной системе, так и др. Основными его функциями можно назвать регуляцию циклов сон-бодрствование, терморегуляцию, влияние на чувства аппетита и насыщения, модуляцию активности макрофагов, усиление секреции ферментов поджелудочной железой, соляной кислоты желудком, сокращение гладкой мускулатуры, выделение гастрина в кишечнике,участие в регуляции дыхания на уровне ствола мозга. GRP стимулирует митотическую активность клеток, в том числе и мелкоклеточного рака легких.
Литоринобладает многими свойствами семейства, в частности способностью к регуляции температуры тела[1, 8].
Кинины
Кинины обладают широким спектром активности и являются связующим звеном в системах регуляции сосудистого тонуса, свертывания крови и фибринолиза. Синтезируются в большинстве тканей, включая ЦНС.
Брадикининсостоит из 9 аминокислотных остатков. Его воздействие приводит к расслаблению гладкой мускулатуры стенок сосудов, бронхов, матки, кишечника. Принимает участие в регуляции гемостаза, электролитного баланса, проницаемости капилляров, местных воспалительных реакций и болевой чувствительности. На мышечные волокна вне сосудов действует противоположным образом, приводя к их констрикции, что имеет значение в развитии воспаления и усиления боли.
Каллидинсостоит из 10 аминокислотных остатков и отличается от брадикинина наличием остатка лизина в начале цепи. По физиологическим эффектам похож на брадикинин [1, 8, 16].
Предшественник – кининоген [20].
Ангиотензины
Ангиотензины I, II, III синтезируются в ЦНС и других тканях и органах. Наиболее изученными функциями данных пептидов является регуляция состояния сердечно-сосудистой системы, водно-солевого обмена и артериального давления. Ангиотензины являются продуктами последовательного гидролитического отщепления с концевой части нескольких аминокислот. Так, из ангиотензина Iобразуется ангиотензин IIи далее с укорочением цепи. Самым мощным из группы является ангиотензин II, который образуется под влиянием ренина и ПДА. Играет роль в формировании механизмов артериальной гипертензии. Входит в состав ренин-ангиотензин-альдостероновой системы. Связан с адренергической системой и тахикининами по механизму действия. Доказано, что ангиотензины принимают участие в процессах обучения, формирования памяти, мотивации, внутреннего подкрепления, болевой чувствительности и контроле эмоций[2, 8].
Предшественник – препротеин ангиотензиноген [20].
Пептиды, кодируемые геном, подобным гену кальцитонина
Кальцитонин (контринсулярный гормон) состоит из 32 аминокислотных остатков. Вырабатывается преимущественно С-клетками щитовидной железы. Кальцитонинподобная иммунореактивность обнаружена в гипофизе, спинномозговой жидкости, легких, тимусе, кишечнике, печени, мочевом пузыре. В мозге наибольшее содержание кальцитонина обнаруживается в зоне, окружающей задний отдел гипоталамуса, срединном возвышении и гипофизе. Принимает активное участие в регуляции водно-солевого баланса. Снижает содержание кальция и фосфора в плазме крови, что приводит к изменению метаболизма и активности клеточных мембран. Оказывает анальгезирующее и аноректическое действие, приводит к вазодилатации, гипотензии, гипергликемии, стимулирует глюконеогенез и гликогенолиз.
Кальцитонин-ген-родственный пептид (CGRP) состоит из 37 аминокислотных остатков. В достаточно больших количествах встречается в центральной и периферической нервной, сердечно-сосудистой, мочеполовой системах, ЖКТ и С-клетках щитовидной железы. Обладает способностью влиять на артериальное давление, в зависимости от сопутствующих факторов приводит к гипо- или гипертензии, является сильным сосудорасширяющим веществом, вызывает тахикардию, участвует в поддержании тонуса коронарных сосудов и модуляции болевой чувствительности, влияет на пищевое поведение и мозговое кровообращение [4, 14].
Предшественник – препро-CALC (кальцитонин) [20].
Атриопептиды
Атриопептиды синтезируются преимущественно в предсердиях. Рецепторы к данной группе пептидов находятся во многих тканях. Наиболее значимыми эффектами являются расслабление гладкой мускулатуры сосудов, что приводит к понижению артериального давления, увеличение проницаемости биологических барьеров и, следовательно, усилению выхода воды в межклеточное пространство, повышению выведения натрия и хлора из-за снижения их реабсорбции в почечных канальцах, что совместно с увеличением клубочковой фильтрации приводит к повышению диуреза, подавление РААС, снижению тонуса мускулатуры кишечника, снижению внутриглазного давления, уменьшению объема ликвора и снижению внутричерепного давления [8].
Предшественник – препро-ANP [20].
Эндозепины
Пептид – ингибитор связывания диазепама (DBI)состоит из большого количества аминокислот. Биологической активностью обладает как сам DBI, так и его фрагменты – эндозепин-6 и октадеканейропептид (6 и 18 аминокислотных остатков в своем составе соответственно). Эти пептиды в большом количестве находятся в ЦНС и являются лигандами бензодиазепиновых рецепторов. По структуре – сильные антагонисты ГАМК, что обусловливает их биологические эффекты. Уровень эндозепинов возрастает в ткани мозга при стрессе и старении. Доказано, что данные пептиды играют роль в формировании реакций на стресс и развитии тревожных состояний – при внутрижелудочковом введении оказывают анксиогенное и проконфликтогенное действие, поэтому поиск их антагонистов может привести к созданию новому виду противотревожных препаратов [5, 8].
Предшественник – BDI в изоформах 1, 2, 3 [20].
Галанин
Галанин состоит из 29 аминокислотных остатков. Рецепторы к нему расположены в гиппокампе, гипоталамусе, миндалине, преоптической зоне, супраоптическом, аркуатном ядрах. Рецепторы подразделяются на 3 типа и все они ассоциированы с G-белком. В экспериментах показано, что при воздействии на рецепторы галанина можнополучить анксиолитический и антидепрессивный эффекты.При совместном воздействии с другими веществами способен оказывать самое разнообразное влияние – при колокализации с ацетилхолином принимает участие в мнестических процессах, доказана роль в развитии болезни Альцгеймера, с норадреналином – формирование социального статуса, с вазопрессином и окситоцином в супраоптическом и паравентрикулярных ядрах приобретает способность влиять на осморегуляцию. Образовавшись в гипоталамусе стимулирует секрецию ЛГ и принимает участие в регуляции обратной связи ГГН-оси. Способен ингибировать секрецию глутамата и электрическую активность аркуатного ядра [8, 14].
Предшественник – препрогаланин [20].
Эндотелины
Эндотелин I, II, III синтезируются преимущественно в эндотелии сосудов, экспрессия их присутствует и в нервной ткани. Являются мощными вазоконстрикторами. Совместно с другими пептидами и гормонами играют важную роль в регуляции состояния эндотелия, развитии почечной ишемии, гипертонии, бронхиальной астмы, сердечной недостаточности и патологиях сосудов мозга. Основным активным пептидом группы является эндотелин I. Эндотелины имеют разных предшественников, экспрессируемых разными генами [8].
Кроме нейропептидов, представленных в данной классификации, существует большое количество соединений, не включенных в нее. На настоящий момент открыто несколько сотен НП, которые по различным причинам невозможно отнести к существующим группам. Приведем некоторые из них.
Орексины
Орексины относятся к группе гипоталамических пептидов и включают в себя орексин А и орексин В, состоящие из 33 и 28 аминокислотных остатков соответственно. Оба пептида взаимодействуют с рецепторами OXR1и OXR2. Нейроны, секретирующие орексины, содержатся в перифорникальной зоне латерального гипоталамического поля. Несмотря на то, что их немного, данные нейроны сильно ветвятся, имея связи с различными отделами мозга, такими как паравентрикулярные, дорсомедиальное аркуатное ядра гипоталамуса, голубое пятно, заднее гипоталамическое поле, спинной мозг. Орексины контролируют потребление пищи, участвуют в регуляции циркадных ритмов и развитии стресс-реакции, половое поведение. Их концентрация в гипоталамусе увеличивается при голодании, однако внутримозговое введение вызывает лишь кратковременное повышение аппетита без заметного влияния на общий объем потребляемой пищи. Нарколепсию и каталепсию связывают с отсутствием орексинов, так как они способны угнетать обе фазы сна и продлевать период бодрствования.Орексины повышают активность симпатической нервной системы и поддерживают мышечный тонус, что имеет значение при усилении физической активности и ее поддержании на высоком уровне. Кроме того, могут положительно влиять на секрецию гормонов ГГН-оси. Предполагается, что орексины могут играть роль в развитии таких заболеваний как хорея Гентингтона, болезнь Паркинсона, синдром обструктивного сонного апноэ, диабета II типа [8, 11, 19].
Лептин
DSIP (дельта-сон-индуцирующий пептид)
DSIP (дельта-сон-индуцирующий пептид) состоит из 9 аминокислотных остатков. До сих пор не выявлен ген, его кодирующий, предшественник и специфические рецепторы с кодирующими их генами. DSIP обнаруживается в нейронах диагональной связки Брока, вентральной перегородки, передних областях гипоталамуса, в областях с гонадотропин-рилизинг-гормонподобной реактивностью, меланинконцентрирующим гормоном, тиреотропным гормоном, пептидами секреторных клеток ЖКТ, меланокортикотропах промежуточной доли гипофиза, стволе мозга, гипофизе и эпифизе; обладает видовыми различиями по локализации. Часто DSIP колокализован с катехоламинами в хромафинных гранулах мозгового слоя надпочечников. При внутрибрюшном введении его крысам, было установлено увеличение в коре больших полушарий содержания норадреналина и серотонина, снижению адреналина без изменения концентрации дофамина, за счет чего он может оказывать тормозное действие на ЦНС. Имеются данные о стимуляции выработки ГАМК и продопаминергическом действии. DSIP блокирует возбуждающее действие глутамата за счет снижения чувствительности «быстрых» ионотропных NMDA-рецепторов глутаматергической системы мозга и последующего снижения глутаматной стрессиндуцированной эксайтотоксичности. Оказывает мембраностабилизирующий эффект на нейрональные, эритроцитарные и лейкоцитарные мембраны. Повышает активность экспрессии гена c-Fos, являющегося маркером активности нейронов, различных отделов лимбической системы, играющих триггерную роль в развитии эмоциональных реакций на стресс и формирующих их нейромедиаторную интеграцию, что приводит к активации соматовегетативных проявлений. Имеет антистрессорное действие и способствует созданию состояния «преадаптации».
Участие DSIP в регуляции циклов сон-бодрствование является спорным, разными авторами приводятся различные данные, от способности подавления до стимуляции фаз сна. Обладает противоэпилептической и антиконвульсантной активностью. Пептид может принимать участие в анальгетических процессах за счет усиления связывания мет-энкефалина с ОР. Усиливает высвобождение мелатонина путем отщепления остатка триптофана с концевой части молекулы. Влияет на снижение активности МАО, подавление синтеза кортиколиберина, стимуляцию синтеза соматотропина, соматостатина и люлиберина. Обусловливает налоксон-зависимую анальгезию. Способен снижать уровень липидов и холестерина, холестеролового коэффициента атерогенности. DSIP способен препятствовать окислительной модификации белков в ходе физиологического старения. Обладает гипергликемической активностью, что может быть связано с увеличением концентрации катехоламинов, снижающих высвобождение инсулина на фоне усиления продукции глюкагона [3, 8].
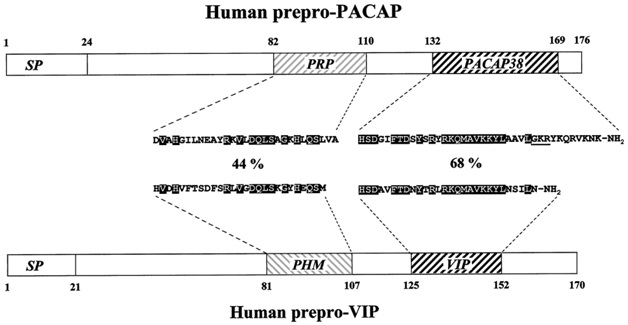
PACAP (пептид, активирующий аденилатциклазу гипофиза)
PACAP (пептид, активирующий аденилатциклазу гипофиза) содержит в своем составе 38 аминокислотных остатков. По структуре и функциям похож на VIP. PACAP широко распространен в мозговых структурах и периферических органах, включая эндокринную систему. Наибольшее количество нейронов, содержащих пептид, находится в гипоталамусе, в частности в супраоптическом и паравентрикулярных ядрах, синтезирующих вазопрессин и окситоцин. Пептид стимулирует их выработку путем активации цАМФ. Может выступать в гипоталамусе в роли нейротрансмиттера и нейромодулятора в регуляции секреции гормонов. Модулирует вазопрессин и окситоцин в регуляции артериального давления и осмоса клеток, принимает участие в модуляции функции мозжечка при физической активности.Предполагается, что PACAP участвует в регуляции ритмичности выработки мелатонина в эпифизе и, соответственно,влияетна циркадные ритмы. Участвует в регуляции пищевого поведения, оказывая анорексигенное действие путем активации цАМФ в супраоптическом и паравентрикулярных ядрах. Имеет важное значение в онтогенезе, ингибируя в развивающемся мозге запрограммированный апоптоз, стимулируя рост нейритов, уменьшая количество митотически делящихся клеток и способствуя дифференцировке нейробластов, оказывает нейропротективное действие при нейротоксичности, вызванной повышенной концентрацией глутамата. Следовательно, в развивающемся мозге PACAP выступает в роли нейротрофического фактора, а в сформировавшемся как нейропротектор. За счет активации аденилатциклазы гипофиза, пептид стимулирует выделение им гормонов [8, 17, 18].
BINP (нейротрофический пептид из поврежденного мозга)
BINP состоит из 13 аминокислотных остатков. Защищает клетки мозга от эксайтотоксичности глутамата, способствует выживанию холинергических нейронов перегородки и дофаминергических нейронов мезэнцефалона в первичной культуре неонатального мозга [8].
Переоценить роль нейропептидов сложно. Их изучение со временем могло бы открыть перспективы для создания лекарств и искусственных регуляторов процессов организма. Сейчас проводится множество исследований и делаются попытки создать препараты на их основе, однако дело осложняется недостаточной изученностью всех свойств нейропептидов и их взаимодействия с другими системами. Как пример таких исследований можно привести следующую таблицу.
Примеры результатов II и III фаз клинических испытаний лигандовнейропептидных рецепторов при психических расстройствах [7].
Подготовила: Жукова С.О.
Редакция : Касьянов Е.Д., помощь в редакции – Anonymous Anonymous


