Глава 7 Генетическая регуляция развития
Генетическая регуляция развития
У природы столько дел в этом мире, ей приходится создавать такую массу разнообразнейших творений, что по временам она и сама не в силах разобраться во всех тех различных процессах, которыми она одновременно занимается.
Уилки Коллинз «Женщина в белом»
Мутации и изменения, происходящие в онтогенезе
Если морфология представляет собой проявление сложного комплекса процессов развития, то эти процессы в свою очередь представляют собой проявление действия целого созвездия генов. Таково важное допущение, положенное в основу этой книги, и мы собрали по крайней мере prima facie данные в пользу того, что у Metazoa часть генома специфически участвует в регуляции онтогенеза и что характер эволюции этой части отличается от характера эволюции структурных генов. До сих пор, однако, мы ограничивались изолированными частными примерами генного контроля морфогенеза и не пытались ответить на центральный вопрос: каким образом гены определяют процесс развития?
Здесь мы в весьма прямом смысле возвращаемся к программе механики развития, выдвинутой Ру (Roux), однако, вместо того чтобы удалять клетки или другие структуры зародыша, с тем чтобы определить их роли в развитии, как это делали представители классической экспериментальной эмбриологии, генетика развития использует мутации как очень тонкий скальпель, дающий возможность уничтожать или изменять отдельные гены.
Генетическая модель, с помощью которой можно изучать любую систему, теоретически находящуюся под генетическим контролем, состоит в следующем. Для того чтобы проанализировать какой-либо процесс, в данном случае онтогенез, исследователь выявляет мутации, изменяющие этот процесс. Выявив такие мутации, он проводит фенотипическое сравнение мутантных особей с нормальными. Это сравнение помогает ему понять, как данный ген влияет на нормальное развитие. Однако, прежде чем продолжить описание метода проведения такого сравнения, следует указать, что воздействие мутаций на онтогенез проявляется двумя основными способами. Это, во-первых, дизруптивные изменения, при которых процесс нормального развития нарушается, что приводит к морфологическим аномалиям (например, к отсутствию некоторых структур). В наиболее резко выраженной форме такие мутации оказываются летальными. Во-вторых, это гомеозисные изменения, при которых под действием мутации развитие отклоняется от нормы, в результате чего какая-либо структура данного организма замещается гомологичным органом или конечностью. Мы отложим дальнейшее обсуждение мутаций этого второго типа до следующей главы и займемся здесь главным образом дизруптивными изменениями.
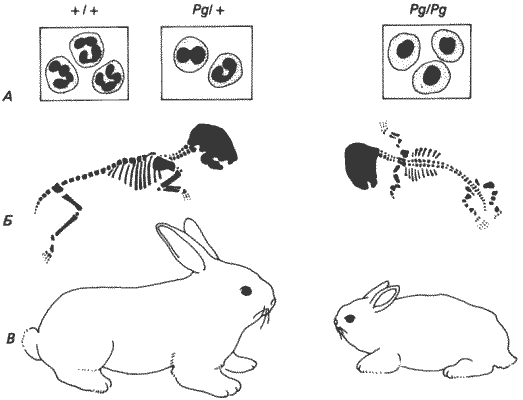
Рис. 7-1. Аномалия Пельгера у кроликов. А. Полиморфноядерные нейтрофилы нормальных ( +/+ ), гетерозиготных (Pg/+) и гомозиготных мутантных (Pg/Pg) кроликов. Б и В. Скелеты и общий вид взрослых кроликов Pg/+ (слева) и Pg/Pg(справа). Обратите внимание на резко выраженную недоразвитость конечностей у гомозиготных особей (Nachtsheim, 1950).
Многочисленные и разнообразные нарушения, вызываемые этой заменой одной аминокислоты, схематически представлены на рис. 7-2. Глядя на этот рисунок, нетрудно понять, что неосведомленный человек, рассматривая схему снизу, может истолковать сам синдром совершенно иначе, чем если бы ему была известна первопричина всех этих нарушений.
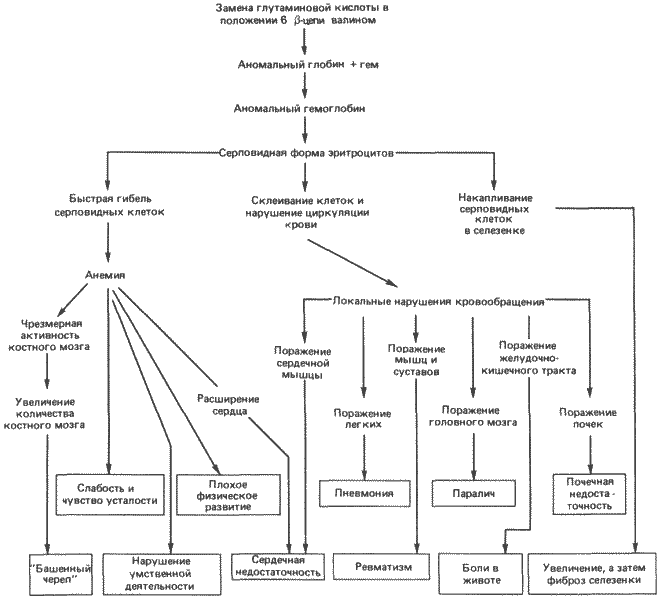
Прежде чем можно будет прийти к каким-то определенным выводам о природе генетических дефектов, необходимо затронуть еще два очень важных момента. Первый из них касается первичного места действия данного гена. Иными словами, существует ли некая специфичная ткань или орган, в которых ген проявляет свою активность? Более того, автономен ли ген в своей активности? Этот вопрос связан с тем, что на самом деле существует плейотропия двух типов: 1) в случае относительной, или зависимой, плейотропии, как при описанной здесь серповидноклеточной анемии, существует одно первичное место действия гена (т. е. эритроциты), а все остальные наблюдаемые нарушения связаны с этим одним дефектом или возникают как его следствие; 2) в случае прямой, или истинной, плейотропии все разнообразные дефекты, возникающие в различных тканях и/или органах, вызываются непосредственным действием одного гена. Грюнберг (Grunberg) на основании своих исследований мутаций у мышей считал преобладающим типом относительную плейотропию. Однако известны также примеры прямых плейотропных эффектов.
Читайте также
Глава XIII Регуляция экспрессии генов
Глава XIII Регуляция экспрессии генов О проблеме регуляции экспрессии генов мы в этой книге говорим фактически во всех главах, рассматривая ее с разных сторон. Существует такое, может быть несколько одностороннее, определение развития: «Понять развитие — это значит
Глава 6. Два энергетических принципа биологического развития
Глава 6. Два энергетических принципа биологического развития В особой земной оболочке, которой является биосфера, характеризуемая необратимыми процессами, жизнь будет увеличивать, а не уменьшать с ходом времени свободную энергию этой оболочки. В. И. Вернадский 6.1.
Глава 11. Генетическая эволюция продуктов поведения животных
Глава 11. Генетическая эволюция продуктов поведения животных Что именно мы понимаем под фенотипическим эффектом гена? Поверхностное знание молекулярной биологии может предложить нам такой ответ: каждый ген кодирует синтез одной полипептидной цепи, и в простейшем смысле
Глава 1. Генетическая информация
Глава 1. Генетическая информация Ключевой проблемой биологии, по-видимому, можно считать вопрос о том, как увековечивает свой опыт живая материя. М. Дельбрюк (1906–1981), американский генетик, лауреат Нобелевской премии 1969 г. При изучении любого биологического феномена
11.5. Генетическая психофизиология
11.5. Генетическая психофизиология Генетическая психофизиология изучает взаимодействие генотипа и факторов среды в формировании физиологических систем организма, обеспечивающих психическую деятельность (Булаева К. Б., 1991).Структурная единица нервной системы – нейрон –
Глава 13. Генетика развития[1]
Глава 13. Генетика развития[1] Пусть не покажется странным, если я позволю себе сказать, что легче понять образование всех небесных тел и причину их движений, короче говоря, происхождение всего современного устройства мироздания, чем точно выяснить на основании механики
Глава 4. Регуляция активности ферментов. Медицинская энзимология
Глава 4. Регуляция активности ферментов. Медицинская энзимология Способы регуляции активности ферментов:1. Изменение количества ферментов.2. Изменение каталитической эффективности фермента.3. Изменение условий протекания реакции.Регуляция количества
Глава 27. Регуляция и взаимосвязь метаболизма
Глава 27. Регуляция и взаимосвязь метаболизма Для нормального функционирования организма должна осуществляться точная регуляция потока метаболитов по анаболическим и катаболическим путям. Все сопутствующие химические процессы должны протекать со скоростями,
ГЛАВА 1 ПЕРИОДЫ РАЗВИТИЯ ЩЕНКОВ
ГЛАВА 1 ПЕРИОДЫ РАЗВИТИЯ ЩЕНКОВ Роды завершают эмбриональный период развития щенка. Послеутробный период развития животных начинается с их рождения и кончается естественной смертью в результате старения.Каждый период развития, состоящий из ряда определенных фаз и
Глава 3. Генетическая информация
Глава 3. Генетическая информация Носителями наследственной информации в природе являются нуклеиновые кислоты. Именно они выполняют три необходимые для жизни функции: хранение, воспроизведение и реализацию этой информации. В ходе эволюции ключевая роль по хранению и
Глава 4. Среда развития
Глава 4. Среда развития Сторожевые пастушьи собаки «К западу от Маньчжурии вплоть до Венгерской равнины простирался океан травы, по его холмистым пространствам кочевники перегоняли свои стада в постоянном поиске пищи. Зимой они укрывались от буранов и метелей в горах,
Глава 10 Адаптации экспрессии генов в процессе развития
Генетическая стоимость развития на молекулярном уровне
Генетическая стоимость развития на молекулярном уровне Попытки перенести оценки числа генов, необходимых для развития дрозофилы, на другие организмы сталкиваются с двумя проблемами. Во-первых, геном дрозофилы, по крайней мере в некоторых существенных отношениях,
5. Теория стадийного развития и особенности развития животных
5. Теория стадийного развития и особенности развития животных В основе управления развитием организмов лежит теория стадийного развития, которую сформулировал академик Т. Д. Лысенко, исходя из работы И. В. Мичурина и многочисленных собственных исследований.Несмотря на
Глава 5. Проблемы индивидуального развития и пол
Глава 5. Проблемы индивидуального развития и пол Генетические аномалии полового развития и половая идентификация В рамках онтогенетической концепции полаДж. Мани полноценный мужчина или женщина должны в своем индивидуальном развитии демонстрировать все восемь
Эпигенетика: теоретические аспекты и практическое значение
В данной статье освещаются современные представления о эпигенетике как новой науке, механизмах эпигенетической регуляции и ее нарушениях, приводящих к развитию заболеваний.
This article highlights the modern views on epigenetics as a new science, the mechanisms of epigenetic regulation and their disorders, leading to the development of diseases.
Эпигенетика — сравнительно недавнее направление биологической науки и пока не так широко известно, как генетика. Под ней понимают раздел генетики, который изучает наследуемые изменения активности генов во время развития организма или деления клеток.
Эпигенетические изменения не сопровождаются перестановкой последовательности нуклеотидов в дезоксирибонуклеиновой кислоте (ДНК).
В организме существуют различные регуляторные элементы в самом геноме, которые контролируют работу генов, в том числе в зависимости от внутренних и внешних факторов. Долгое время эпигенетику не признавали, т. к. было мало информации о природе эпигенетических сигналов и механизмах их реализации.
Структура генома человека
В 2002 г. в результате многолетних усилий большого числа ученых разных стран закончена расшифровка строения наследственного аппарата человека, который заключен в главной молекуле ДНК. Это одно из выдающихся достижений биологии начала ХХI века.
ДНК, в которой находится вся наследственная информация о данном организме, называется геномом. Гены — это отдельные участки, занимающие очень небольшую часть генома, но при этом составляют его основу. Каждый ген отвечает за передачу в организме человека данных о строении рибонуклеиновой кислоты (РНК) и белка. Структуры, которые передают наследственную информацию, называют кодирующими последовательностями. В результате проекта «Геном» были получены данные, согласно которым геном человека оценивался в более чем 30 000 генов. В настоящее время, в связи с появлением новых результатов масс-спектрометрии, геном предположительно насчитывает около 19 000 генов [1].
Генетическая информация каждого человека содержится в ядре клетки и расположена в особых структурах, получивших название хромосомы. Каждая соматическая клетка содержит два полных набора (диплоидный) хромосом. В каждом единичном наборе (гаплоидном) присутствует 23 хромосомы — 22 обычные (аутосомы) и по одной половой хромосоме — Х или Y.
Молекулы ДНК, содержащиеся во всех хромосомах каждой клетки человека, представляют собой две полимерные цепи, закрученные в правильную двойную спираль.
Обе цепи удерживают друг друга четырьмя основаниями: аденин (А), цитозин (Ц), гуанин (Г) и тиамин (Т). Причем основание А на одной цепочке может соединиться только с основанием Т на другой цепочке и аналогично основание Г может соединяться с основанием Ц. Это называется принципом спаривания оснований. При других вариантах спаривание нарушает всю целостность ДНК.
ДНК существует в виде тесного комплекса со специализированными белками, и вместе они составляют хроматин.
Гистоны — это нуклеопротеины, основная составляющая хроматина. Им свойственно образование новых веществ путем присоединения двух структурных элементов в комплекс (димер), что является особенностью для последующей эпигенетической модификации и регуляции.
ДНК, хранящая генетическую информацию, при каждом клеточном делении самовоспроизводится (удваивается), т. е. снимает с самой себя точные копии (репликация). Во время клеточного деления связи между двумя цепями двойной спирали ДНК разрушаются и нити спирали разделяются. Затем на каждой из них строится дочерняя цепь ДНК. В результате молекула ДНК удваивается, образуются дочерние клетки.
ДНК служит матрицей, на которой происходит синтез разных РНК (транскрипция). Этот процесс (репликация и транскрипция) осуществляется в ядрах клеток, а начинается он с области гена, называемой промотором, на котором связываются белковые комплексы, копирующие ДНК для формирования матричной РНК (мРНК).
В свою очередь последняя служит не только носителем ДНК-информации, но и переносчиком этой информации для синтеза белковых молекул на рибосомах (процесс трансляции).
В настоящее время известно, что зоны гена человека, кодирующие белки (экзоны), занимают лишь 1,5% генома [2]. Большая часть генома не имеет отношения к генам и инертна в плане передачи информации. Выявленные зоны гена, не кодирующие белки, называются интронами.
Первая копия мРНК, полученная с ДНК, содержит в себе весь набор экзонов и интронов. После этого специализированные белковые комплексы удаляют все последовательности интронов и соединяют друг с другом экзоны. Этот процесс редактирования называется сплайсингом.
Эпигенетика объясняет один из механизмов, с помощью которого клетка способна контролировать синтез производимого ею белка, определяя в первую очередь, сколько копий мРНК можно получить с ДНК.
Итак, геном — это не застывшая часть ДНК, а динамическая структура, хранилище информации, которую нельзя свести к одним генам.
Развитие и функционирование отдельных клеток и организма в целом не запрограммированы автоматически в одном геноме, но зависят от множества различных внутренних и внешних факторов. По мере накопления знаний выясняется, что в самом геноме существуют множественные регуляторные элементы, которые контролируют работу генов. Сейчас это находит подтверждение во множестве экспериментальных исследований на животных [3].
При делении во время митоза дочерние клетки могут наследовать от родительских не только прямую генетическую информацию в виде новой копии всех генов, но и определенный уровень их активности. Такой тип наследования генетической информации получил название эпигенетического наследования.
Эпигенетические механизмы регуляции генов
Предметом эпигенетики является изучение наследования активности генов, не связанной с изменением первичной структуры входящей в их состав ДНК. Эпигенетические изменения направлены на адаптацию организма к изменяющимся условиям его существования.
Впервые термин «эпигенетика» предложил английский генетик Waddington в 1942 г. Разница между генетическими и эпигенетическими механизмами наследования заключается в стабильности и воспроизводимости эффектов [4].
Генетические признаки фиксируются неограниченное число, пока в гене не возникает мутация. Эпигенетические модификации обычно отображаются в клетках в пределах жизни одного поколения организма. Когда данные изменения передаются следующим поколениям, то они могут воспроизводиться в 3–4 генерациях, а затем, если стимулирующий фактор пропадает, эти преобразования исчезают.
Молекулярная основа эпигенетики характеризуется модификацией генетического аппарата, т. е. активации и репрессии генов, не затрагивающих первичную последовательность нуклеотидов ДНК.
Эпигенетическая регуляция генов осуществляется на уровне траскрипции (время и характер транскрипции гена), при отборе зрелых мРНК для транспорта их в цитоплазму, при селекции мРНК в цитоплазме для трансляции на рибосомах, дестабилизации определенных типов мРНК в цитоплазме, избирательной активации, инактивации молекул белков после их синтеза.
Совокупность эпигенетических маркеров представляет собой эпигеном. Эпигенетические преобразования могут влиять на фенотип.
Эпигенетика играет важную роль в функционировании здоровых клеток, обеспечивая активацию и репрессию генов, в контроле транспозонов, т. е. участков ДНК, способных перемещаться внутри генома, а также в обмене генетического материала в хромосомах [5].
Эпигенетические механизмы участвуют в геномном импритинге (отпечаток) — процессе, при котором экспрессия определенных генов осуществляется в зависимости от того, от какого родителя поступили аллели. Импритинг реализуется через процесс метилирования ДНК в промоторах, в результате чего транскрипция гена блокируется.
Эпигенетические механизмы обеспечивают запуск процессов в хроматине через модификации гистонов и метилирование ДНК. За последние два десятилетия существенно изменились представления о механизмах регуляции транскрипции эукариот. Классическая модель предполагала, что уровень экспрессии определяется транскрипционными факторами, связывающимися с регуляторными областями гена, которые инициируют синтез матричной РНК. Гистонам и негистоновым белкам отводилась роль пассивной упаковочной структуры для обеспечения компактной укладки ДНК в ядре.
В последующих исследованиях была показана роль гистонов в регуляции трансляции. Был обнаружен так называемый гистоновый код, т. е. модификация гистонов, неодинаковая в разных районах генома. Видоизмененные гистоновые коды могут приводить к активизации и репрессии генов [6].
Модификациям подвергаются различные части структуры генома. К концевым остаткам могут присоединяться метильные, ацетильные, фосфатные группы и более крупные белковые молекулы.
Все модификации являются обратимыми и для каждой существуют ферменты, которые ее устанавливают или удаляют.
Метилирование ДНК
У млекопитающих метилирование ДНК (эпигенетический механизм) было изучено раньше других. Показано, что он коррелирует с репрессией генов. Экспериментальные данные показывают, что метилирование ДНК является защитным механизмом, подавляющим значительную часть генома чужеродной природы (вирусы и др.).
Метилирование ДНК в клетке контролирует все генетические процессы: репликацию, репарацию, рекомбинацию, транскрипцию, инактивацию Х-хромосомы. Метильные группы нарушают ДНК-белковое взаимодействие, препятствуя связыванию транскрипционных факторов. Метилирование ДНК влияет на структуру хроматина, блокирует транскрипционные репрессоры [7].
Действительно, повышение уровня метилирования ДНК коррелирует с относительным увеличением содержания некодирующей и повторяющейся ДНК в геномах высших эукариот. Экспериментальные данные показывают, что это происходит потому, что метилирование ДНК служит главным образом как защитный механизм, чтобы подавлять значительную часть генома чужеродного происхождения (реплицированные перемещающиеся элементы, вирусные последовательности, другие повторяющиеся последовательности).
Профиль метилирования — активирование или угнетение — меняется в зависимости от средовых факторов. Влияние метилирования ДНК на структуру хроматина имеет большое значение для развития и функционирования здорового организма, чтобы подавлять значительную часть генома чужеродного происхождения, т. е. реплицированные перемещающиеся элементы, вирусные и другие повторяющиеся последовательности.
Метилирование ДНК происходит путем обратимой химической реакции азотистого основания — цитозина, в результате чего метильная группа СН3 присоединяется к углероду с образованием метилцитозина. Этот процесс катализируется ферментами ДНК-метилтрансферазами. Для метилирования цитозина необходим гуанин, в результате образуется два нуклеотида, разделенные фосфатом (СрG).
Скопление неактивных последовательностей СрG называется островками СрG. Последние представлены в геноме неравномерно [8]. Большинство из них выявляются в промоторах генов. Метилирование ДНК происходит в промоторах генов, в транскрибируемых участках, а также в межгенных пространствах.
Гиперметилированные островки вызывают инактивацию гена, что нарушает взаимодействие регуляторных белков с промоторами.
Метилирование ДНК оказывает огромное влияние на экспрессию генов и, в конечном счете, на функцию клеток, тканей и организма в целом. Установлена прямая зависимость между высоким уровнем метилирования ДНК и количеством репрессированных генов.
Удаление метильных групп из ДНК в результате отсутствия метилазной активности (пассивное деметилирование) реализуется после репликации ДНК. При активном деметилировании участвует ферментативная система, превращающая 5-метилцитозин в цитозин независимо от репликации. Профиль метилирования меняется в зависимости от средовых факторов, в которых находится клетка.
Утрата способности поддерживать метилирование ДНК может приводить к иммунодефициту, злокачественным опухолям и другим заболеваниям [9].
Долгое время механизм и ферменты, вовлеченные в процесс активного деметилирования ДНК, оставались неизвестными.
Ацетилирование гистонов
Существует большое число посттрансляционных модификаций гистонов, которые формируют хроматин. В 1960-е годы Винсент Олфри идентифицировал ацетилирование и фосфорилирование гистонов из многих эукариот [8].
Ферменты ацетилирования и деацетилирования (ацетилтрансферазы) гистонов играют роль в ходе транскрипции. Эти ферменты катализируют ацетилирование локальных гистонов. Деацетилазы гистонов репрессируют транскрипцию.
Эффект ацетилирования это ослабление связи между ДНК и гистонами из-за изменения заряда, в результате чего хроматин становится доступным для факторов транскрипции.
Ацетилирование представляет собой присоединение химической ацетил-группы (аминокислоты лизин) на свободный участок гистона. Как и метилирование ДНК, ацетилирование лизина представляет собой эпигенетический механизм для изменения экспрессии генов, не влияющих на исходную последовательность генов. Шаблон, по которому происходят модификации ядерных белков, стали называть гистоновым кодом.
Гистоновые модификации принципиально отличаются от метилирования ДНК. Метилирование ДНК представляет собой очень стабильное эпигенетическое вмешательство, которое чаще закрепляется в большинстве случаев.
Подавляющее большинство гистоновых модификаций более вариативно. Они влияют на регуляцию экспрессии генов, поддержание структуры хроматина, дифференциацию клеток, канцерогенез, развитие генетических заболеваний, старение, репарацию ДНК, репликацию, трансляцию. Если гистоновые модификации идут на пользу клетки, то они могут продолжаться довольно долго [8].
Одним из механизмов взаимодействия между цитоплазмой и ядром является фосфорилирование и/или дефосфорилирование транскрипционных факторов. Гистоны были одними из первых белков, фосфорилирование которых было обнаружено. Это осуществляется с помощью протеинкиназ.
Под контролем фосфорилируемых транскрипционных факторов находятся гены, в том числе гены, регулирующие пролиферацию клеток. При подобных модификациях в молекулах хромосомных белков происходят структурные изменения, которые приводят к функциональным изменениям хроматина.
Помимо описанных выше посттрансляционных модификаций гистонов имеются более крупные белки, такие как убиквитин, SUMO и др., которые могут присоединяться с помощью ковалентной связи к боковым аминогруппам белка-мишени, оказывая воздействие на их активность.
Эпигенетические изменения могут передаваться по наследству (трансгенеративная эпигенетическая наследственность). Однако в отличие от генетической информации, эпигенетические изменения могут воспроизводиться в 3–4 поколениях, а при отсутствии фактора, стимулирующего эти изменения, исчезают. Передача эпигенетической информации происходит в процессе мейоза (деления ядра клетки с уменьшением числа хромосом вдвое) или митоза (деления клеток).
Модификации гистонов играют фундаментальную роль в нормальных процессах и при заболеваниях.
Регуляторные РНК
Молекулы РНК выполняют в клетке множество функций. Одной из них является регуляция экспрессии генов. За эту функцию отвечают регуляторные РНК, к которым относятся антисмысловые РНК (aRNA), микроРНК (miRNA) и малые интерферирующие РНК (siRNA)
Механизм действия разных регуляторных РНК схож и заключается в подавлении экспрессии генов, реализующейся путем комплементарного присоединения регуляторной РНК к мРНК, с образованием двухцепочечной молекулы (дцРНК). Само по себе образование дцРНК приводит к нарушению связывания мРНК с рибосомой или другими регуляторными факторами, подавляя трансляцию. Также после образования дуплекса возможно проявление феномена РНК-интерференции — фермент Dicer, обнаружив в клетке двухцепочечную РНК, «разрезает» ее на фрагменты. Одна из цепей такого фрагмента (siRNA) связывается комплексом белков RISC (RNA-induced silencing complex) [10].
В результате деятельности RISC одноцепочечный фрагмент РНК соединяется с комплементарной последовательностью молекулы мРНК и вызывает разрезание мРНК белком семейства Argonaute. Данные события приводят к подавлению экспрессии соответствующего гена.
Физиологические функции регуляторных РНК разнообразны — они выступают основными небелковыми регуляторами онтогенеза, дополняют «классическую» схему регуляции генов.
Геномный импритинг
Человек обладает двумя копиями каждого гена, один из которых унаследован от матери, другой от отца. Обе копии каждого гена имеют возможность быть активной в любой клетке. Геномный импритинг это эпигенетически избирательная экспрессия только одного из аллельных генов, наследуемых от родителей. Геномный импритинг затрагивает и мужское и женское потомство. Так, импритингованный ген, активный на материнской хромосоме, будет активным на материнской хромосоме и «молчащим» на отцовской у всех детей мужского и женского пола. Гены, подверженные геномному импритингу, в основном кодируют факторы, регулирующие эмбриональный и неонатальный рост [11].
Импритинг представляет сложную систему, которая может ломаться. Импритинг наблюдается у многих больных с хромосомными делециями (утраты части хромосом). Известны заболевания, которые у человека возникают в связи с нарушением функционирования механизма импритинга.
Прионы
В последние десятилетие внимание привлечено к прионам, белкам, которые могут вызывать наследуемые фенотипические изменения, не изменяя нуклеотидной последовательности ДНК. У млекопитающих прионный белок расположен на поверхности клеток. При определенных условиях нормальная форма прионов может изменяться, что модулирует активность этого белка.
Викнер выразил уверенность в том, что этот класс белков является одним из многих, которые составляют новую группу эпигенетических механизмов, требующих дальнейшего изучения. Он может находиться в нормальном состоянии, а в измененном состоянии прионные белки могут распространяться, т. е. стать инфекционными [12].
Первоначально прионы были открыты как инфекционные агенты нового типа, но сейчас считают, что они представляют собой феномен общебиологический и являются носителями информации нового типа, хранимой в конформации белка. Феномен прионов лежит в основе эпигенетической наследственности и регуляции экспрессии генов на посттрансляционном уровне.
Эпигенетика в практической медицине
Эпигенетические модификации контролируют все стадии развития и функциональную активность клеток. Нарушение механизмов эпигенетической регуляции напрямую или косвенно связано с множеством заболеваний.
К заболеваниям с эпигенетической этиологией относят болезни импринтинга, которые в свою очередь делятся на генные и хромосомные, всего в настоящее время насчитывают 24 нозологии.
При болезнях генного импринтинга наблюдается моноаллельная экспрессия в локусах хромосом одного из родителей. Причиной являются точечные мутации в генах, дифференцированно экспрессирующихся в зависимости от материнского и отцовского происхождения и приводящих к специфическому метилированию цитозиновых оснований в молекуле ДНК. К ним относят: синдром Прадера–Вилли (делеция в отцовской хромосоме 15) — проявляется черепно-лицевым дисморфизмом, низким ростом, ожирением, мышечной гипотонией, гипогонадизмом, гипопигментацией и задержкой умственного развития; синдром Ангельмана (делеция критического района, находящегося в 15-й материнской хромосоме), основными признаками которого являются микробрахицефалия, увеличенная нижняя челюсть, выступающий язык, макростомия, редкие зубы, гипопигментация; синдром Беквитта–Видемана (нарушение метилирования в коротком плече 11-й хромосомы), проявляющийся классической триадой, включающей макросомию, омфалоцеле макроглоссию и др. [13].
К числу важнейших факторов, влияющих на эпигеном, относятся питание, физическая активность, токсины, вирусы, ионизирующая радиация и др. Особенно чувствительным периодом к изменению эпигенома является внутриутробный период (особенно охватывающий два месяца после зачатия) и первые три месяца после рождения. В период раннего эмбриогенеза геном удаляет большую часть эпигенетических модификаций, полученных от предыдущих поколений. Но процесс репрограммирования продолжается в течение всей жизни [14].
К заболеваниям, где нарушение генной регуляции является частью патогенеза, можно отнести некоторые виды опухолей, сахарный диабет, ожирение, бронхиальную астму, различные дегенеративные и другие болезни [15, 16].
Эпигоном при раке характеризуется глобальными изменениями в метилировании ДНК, модификации гистонов, а также изменением профиля экспрессии хроматин-модифицирующих ферментов.
Опухолевые процессы характеризуются инактивацией посредством гиперметилирования ключевых генов-супрессоров и посредством гипометилирования активацией целого ряда онкогенов, факторов роста (IGF2, TGF) и мобильных повторяющихся элементов, расположенных в районах гетерохроматина [17].
Так, в 19% случаев гипернефроидные опухоли почки ДНК островков СрG была гиперметилированной, а при раке груди и немелкоклеточной карциноме легких выявлена взаимосвязь между уровнями гистонового ацетилирования и экспрессией супрессора новообразований — чем ниже уровни ацетилирования, тем слабее экспрессия гена.
В настоящее время уже разработаны и внедрены в практику противоопухолевые лекарственные препараты, основанные на подавлении активности ДНК-метилтрансфераз, что приводит к снижению метилирования ДНК, активации генов-супрессоров опухолевого роста и замедлению пролиферации опухолевых клеток. Так, для лечения миелодиспластического синдрома в комплексной терапии применяют препараты децитабин (Decitabine) и азацитидин (Azacitidine) [18]. С 2015 г. для лечения множественной миеломы в сочетании с классической химиотерапией применяют панобиностат (Panibinostat), являющийся ингибитором гистоновой деацитилазы [19]. Данные препараты по данным клинических исследований оказывают выраженный положительный эффект на уровень выживаемости и качество жизни пациентов.
Изменения экспрессии тех или иных генов могут происходить и в результате действия на клетку факторов внешней среды. В развитии сахарного диабета 2-го типа и ожирения играет роль так называемая «гипотеза экономного фенотипа», согласно которой недостаток питательных веществ в процессе эмбрионального развития приводит к развитию патологического фенотипа [20]. На моделях животных был выявлен участок ДНК (локус Pdx1), в котором под влиянием недостаточности питания снижался уровень ацетилирования гистонов, при этом наблюдались замедление деления и нарушения дифференцировки B-клеток островков Лангерганса и развития состояния, схожего с сахарным диабетом 2-го типа [21].
Активно развиваются и диагностические возможности эпигенетики. Появляются новые технологии, способные анализировать эпигенетические изменения (уровень метилирования ДНК, экспрессию микроРНК, посттрансляционные модификации гистонов и др.), такие как иммунопреципитация хроматина (CHIP), проточная цитометрия и лазерное сканирование, что дает основания полагать, что в ближайшее время будут выявлены биомаркеры для изучения нейродегенеративных заболеваний, редких, многофакторных болезней и злокачественных новообразований и внедрены в качестве методов лабораторной диагностики [22].
Итак, в настоящее время эпигенетика бурно развивается. С ней связывают прогресс в биологии и медицине.
Литература
ФГБОУ ВО РНИМУ им. Н. И. Пирогова МЗ РФ, Москва


